Aminoacil-tRNA Sintetasi
Le aminoacil-tRNA sintetasi sono una famiglia di enzimi responsabili del caricamento (amminoacilazione) degli amminoacidi sui corrispondenti tRNA.
Questo processo è caratterizzato da un’estrema precisione, essenziale per mantenere l’accuratezza dell’espressione genica complessiva.

Assenza di Controllo Ribosomiale (Esperimento di Chapeville)
L’estrema accuratezza delle aminoacil-tRNA sintetasi è critica perché il ribosoma non verifica l’identità dell’amminoacido portato dal tRNA, ma controlla unicamente la corrispondenza e l’appaiamento codone-anticodone. Se un tRNA viene caricato con un amminoacido errato, quest’ultimo verrà incorporato nella catena polipeptidica senza alcun blocco.
Questa assenza di controllo è stata dimostrata dall’esperimento di Chapeville et al.:
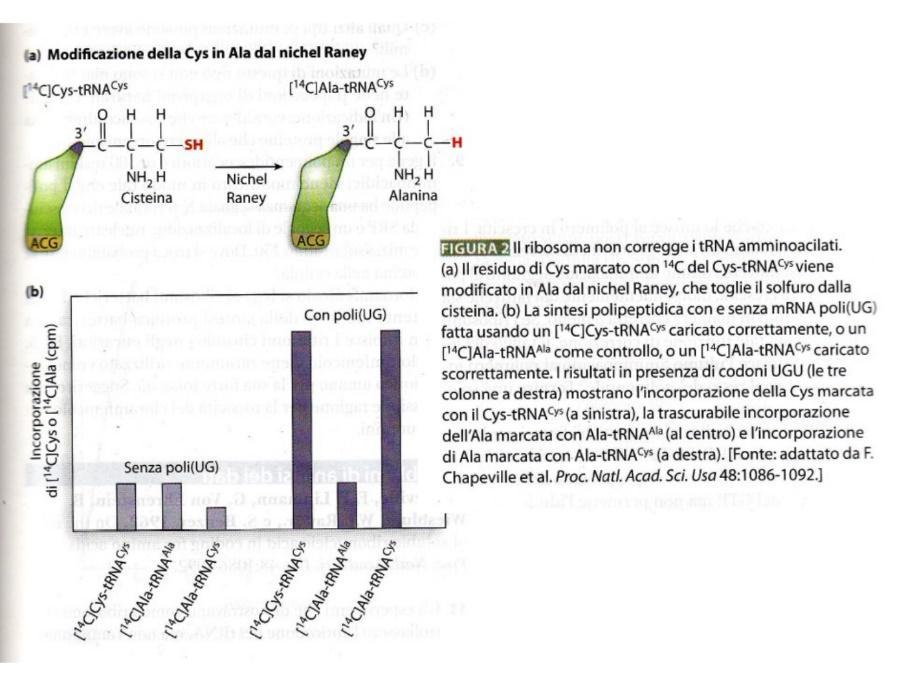
- Caricamento Specifico: Il tRNA per la cisteina () è stato caricato con cisteina radioattiva (-Cisteina).
- Modificazione Chimica: Una porzione di questo complesso è stata trattata chimicamente (tramite riduzione con nichel Raney) per convertire la cisteina in alanina senza distaccarla dal tRNA, ottenendo un caricato con alanina radioattiva (-Alanina).
- Traduzione in vitro: I complessi sono stati saggiati in un sistema di traduzione in vitro con un polinucleotide sintetico contenente solo residui di cisteina ():
- Provette con caricato con cisteina Sintesi proteica elevata (radioattività incorporata).
- Provette con (tRNA dell’alanina) caricato con alanina Nessuna sintesi (il ribosoma rifiuta l’anticodone non complementare).
- Provette con caricato con alanina Sintesi proteica elevata identica al controllo positivo. Il ribosoma non rileva che l’amminoacido legato al tRNA è alanina anziché cisteina, incorporandola erroneamente nella proteina.
Meccanismo di Reazione (Amminoacilazione)
Esistono 20 aminoacil-tRNA sintetasi distinte nella cellula, una per ciascun amminoacido. Il caricamento avviene in due passaggi sequenziali che richiedono il consumo di una molecola di ATP:
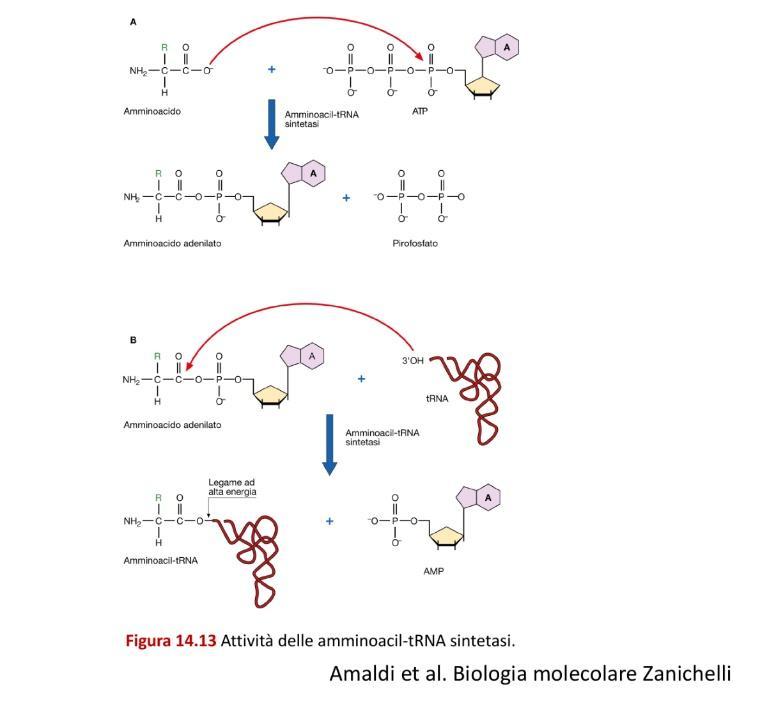
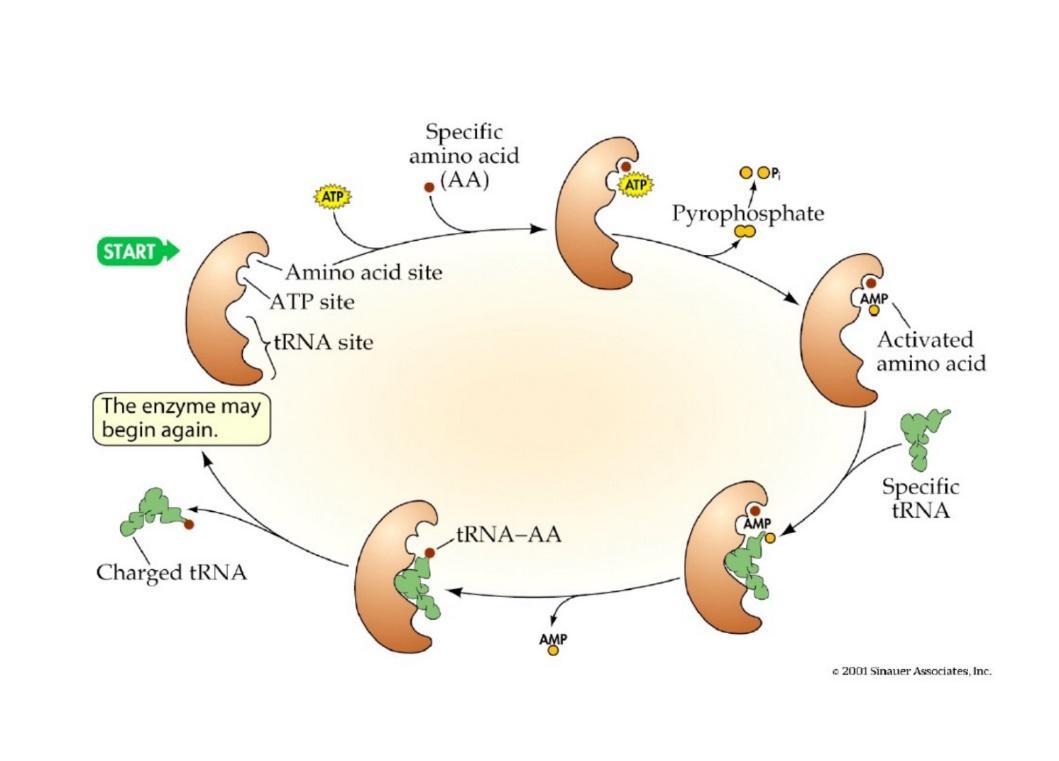
1. Attivazione dell’Amminoacido (Adenilazione)
L’amminoacido viene legato all’ATP nel sito attivo dell’enzima. Si forma un legame anidridico ad alta energia (anidride mista: il gruppo carbossilico dell’amminoacido condivide un ossigeno con il fosfato α dell’AMP, con perdita d’acqua), con rilascio e idrolisi di pirofosfato ():
2. Trasferimento al tRNA
L’amminoacil-AMP attivato rimane legato all’enzima. All’arrivo del tRNA specifico, l’energia del legame ad alta energia dell’amminoacil-AMP viene sfruttata per trasferire l’amminoacido all’ossidrile in 3’ dell’adenosina terminale (della sequenza CCA) del tRNA, formando un legame estere ad alta energia e liberando AMP: Una volta completato il caricamento, il cambiamento conformazionale dell’enzima determina il rilascio del tRNA carico (aminoacil-tRNA), rendendo l’enzima pronto per un nuovo ciclo.
Meccanismi di Riconoscimento e Specificità
Per evitare errori (attacco di amminoacidi errati), l’enzima possiede tre tasche di legame specifiche (per amminoacido, ATP e tRNA):
- Specificità per l’Amminoacido: Il sito attivo dell’enzima è complementare per forma, dimensione, carica elettrica e idrofobicità al gruppo radicale () dell’amminoacido.
- La discriminazione tra amminoacidi molto diversi (es. glicina rispetto a triptofano) è favorita dalle differenze di ingombro sterico.
- Specificità per il tRNA: L’enzima riconosce il tRNA specifico interagendo non solo con la sequenza dell’anticodone sul braccio dell’anticodone, ma anche con elementi strutturali distribuiti lungo gli altri bracci. La forma e dimensione globale del tRNA non sono sufficienti per il riconoscimento, in quanto uguali in tutti i tRNA.
(Sezione espansa con: sbobina 25)
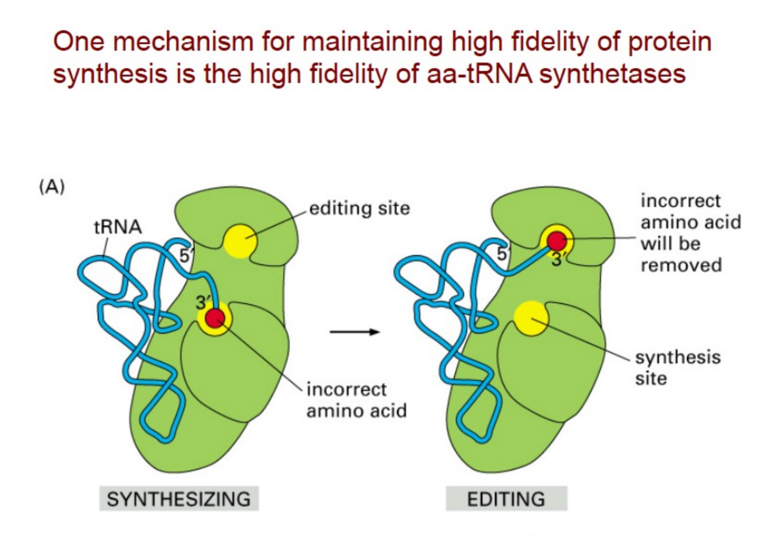
Setaccio Molecolare (Editing)
Il riconoscimento dell’amminoacido corretto si basa su interazioni deboli (forze di van der Waals, legami idrogeno, idrofilia/idrofobicità) e sull’ingombro sterico. Sebbene una tasca di legame piccola impedisca l’ingresso di amminoacidi più grandi, essa non può escludere l’ingresso di amminoacidi più piccoli aventi affinità chimica simile. In questi casi, per evitare errori di incorporazione, l’aminoacil-tRNA sintetasi sfrutta un meccanismo di correzione di bozze (editing) noto come setaccio molecolare:
- Frequenza di errore: Grazie a questo sistema, l’accuratezza finale del caricamento raggiunge un tasso di errore di solo un errore ogni 10.000 amminoacidi.
- Sito di Editing (doppio setaccio): L’amminoacido errato viene deviato in un secondo sito attivo dell’enzima, il sito di editing, dove viene idrolizzato dal tRNA ed espulso a spese di un GTP (così riportato a lezione; molti manuali descrivono l’editing idrolitico senza consumo di GTP — il costo energetico principale è l’AMP già speso nell’attivazione).
- Il meccanismo è a due setacci: il primo setaccio (sito di sintesi) esclude per ingombro gli amminoacidi più grandi di quello corretto; il secondo setaccio (sito di editing) cattura e idrolizza quelli più piccoli o simili che sono comunque riusciti a entrare. Il sito di editing è dimensionato per accogliere l’errore, non l’amminoacido giusto.

Esempi di Selezionamento e Discriminazione
- Fenilalanina (Phe) vs Tirosina (Tyr): La tirosina si distingue dalla fenilalanina per la presenza di un gruppo ossidrile () sull’anello aromatico. Questo rende la tirosina leggermente più ingombrante e capace di formare un legame a idrogeno aggiuntivo.
- Isoleucina (Ile) vs Valina (Val): L’isoleucina possiede un gruppo metilico aggiuntivo rispetto alla valina (isopropile in valina, sec-butile in isoleucina).
- La valina può accidentalmente entrare nella tasca dell’Isoleucil-tRNA sintetasi lasciando uno spazio vuoto dovuto al gruppo metile mancante.
- Questo determina interazioni deboli meno stabili, riducendo la vita media del complesso e provocando lo scivolamento di Val-AMP verso il sito di editing, dove viene occupato temporaneamente e infine idrolizzato.
- Al contrario, l’isoleucina non può entrare nella Valil-tRNA sintetasi a causa del proprio eccessivo ingombro sterico.
🔗 Collegamenti
- Sintesi Proteica — ⬆️ causa
- tRNA — 🔗 stesso meccanismo / stessa via
- Enzimi — 📋 fa parte di
- Maturazione del tRNA — 🔗 stesso meccanismo / stessa via