Promotore
Il promotore è una sequenza specifica di DNA situata a monte della porzione codificante di un gene, responsabile del legame dell’RNA polimerasi e della corretta impostazione dell’inizio della trascrizione.
Scoperta Sperimentale (Esperimento di Pribnow)
La struttura del promotore batterico è stata chiarita da David Pribnow tramite esperimenti di digestione enzimatica:
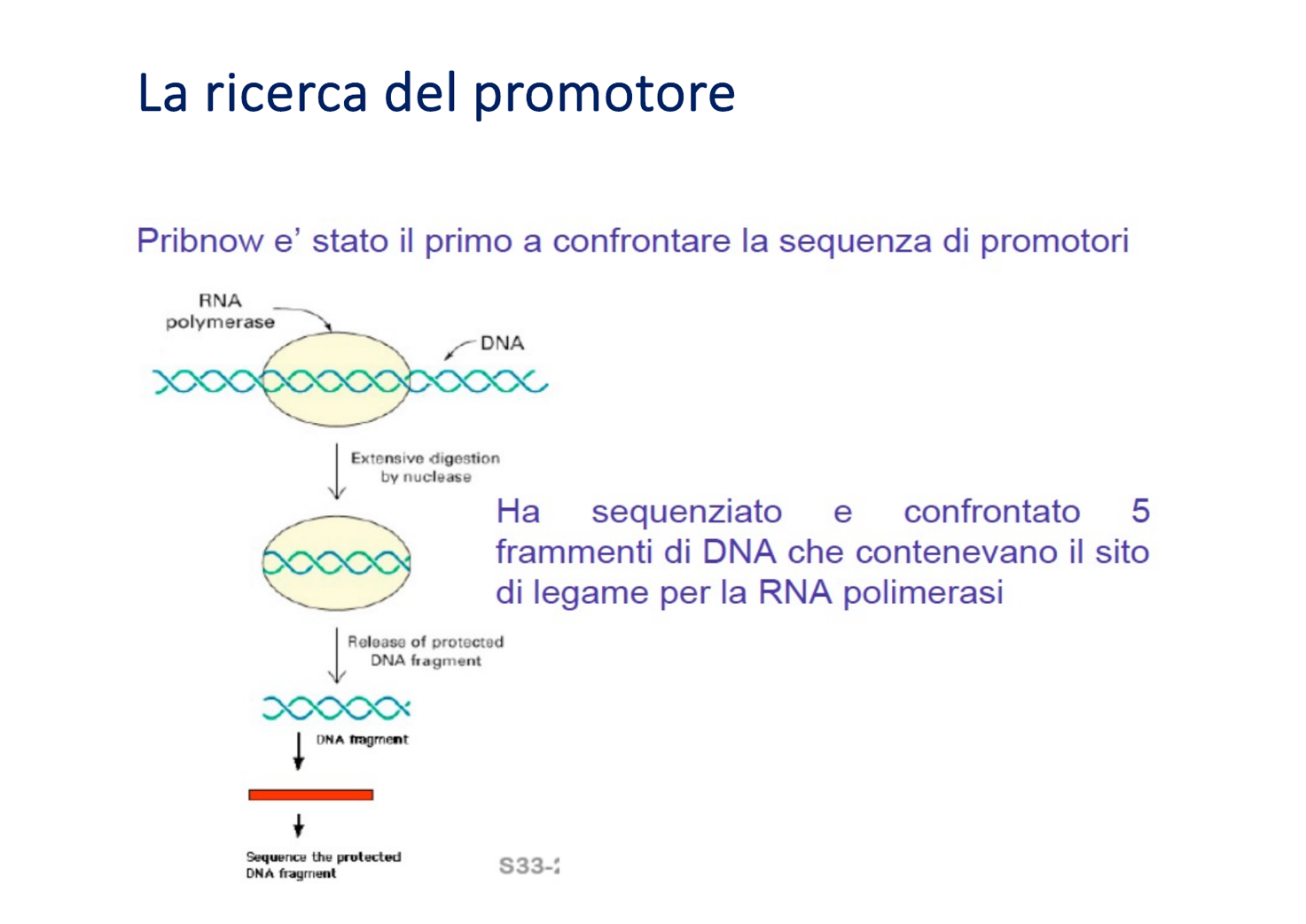
- Ha incubato molecole di oloenzima con il genoma di E. coli, permettendo il legame stabile sul DNA (complesso chiuso).
- Ha aggiunto esonucleasi per digerire tutto il DNA eccetto i segmenti protetti dall’ingombro sterico dell’oloenzima.
- Ha isolato e sequenziato i frammenti protetti rimasti intatti.
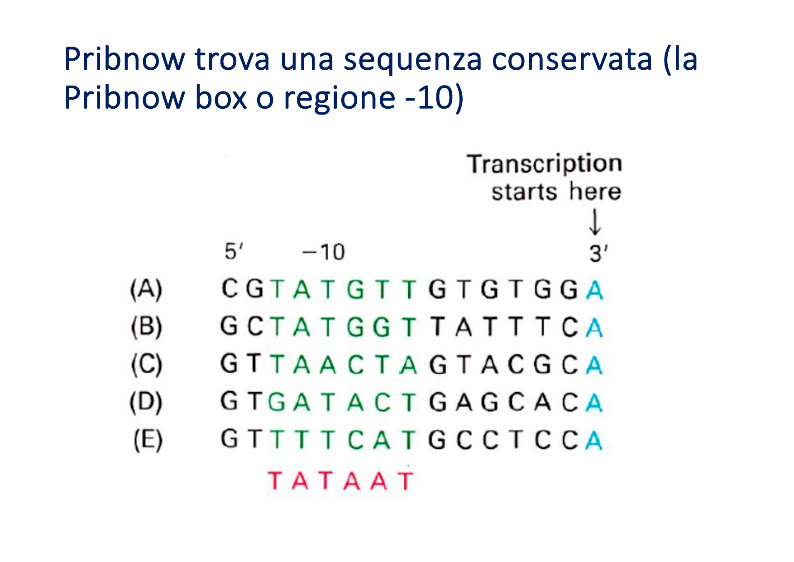
- Risultati: Contrariamente all’ipotesi iniziale di sequenze identiche per tutti i promotori, Pribnow scoprì che le sequenze erano in gran parte diverse, ma condividevano brevi tratti altamente conservati.
Anatomia del Promotore Batterico
Grazie al sequenziamento comparativo di centinaia di promotori di E. coli, è stato definito il modello strutturale standard del promotore procariotico:

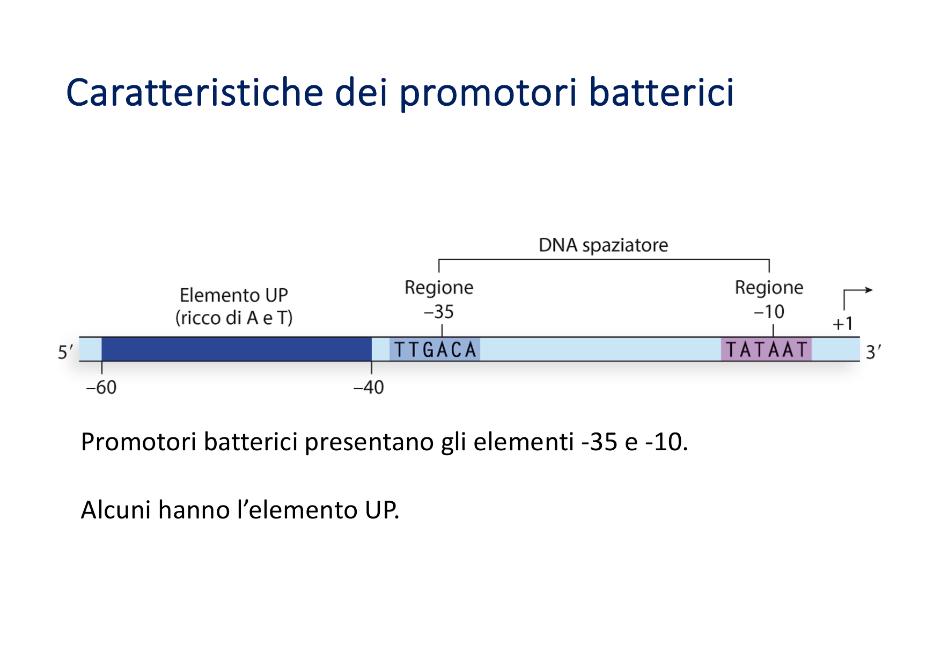
- Sito d’inizio (+1): Il primo nucleotide a essere sintetizzato, che nella grande maggioranza dei casi è una purina (Adenina o Guanina).
- Regione -10 (Pribnow Box): Sequenza consenso di 6 nucleotidi TATAAT, centrata a circa 10 paia di basi a monte del sito d’inizio. (Nota: non va confusa con la TATA box, che è un elemento promotore esclusivo degli eucarioti).
- Regione -35: Sequenza consenso di 6 nucleotidi TTGACA, situata a circa 35 paia di basi a monte del sito d’inizio.
- Regione interposta (Spacer): La sequenza di DNA che separa la regione -10 dalla regione -35. Per consentire il corretto allineamento e aggancio spaziale dell’RNA polimerasi, deve avere una lunghezza ottimale compresa tra 16 e 19 paia di basi.
Funzioni delle Regioni -35 e -10
Esperimenti di mutagenesi guidata hanno chiarito il ruolo funzionale delle due regioni consenso del promotore:

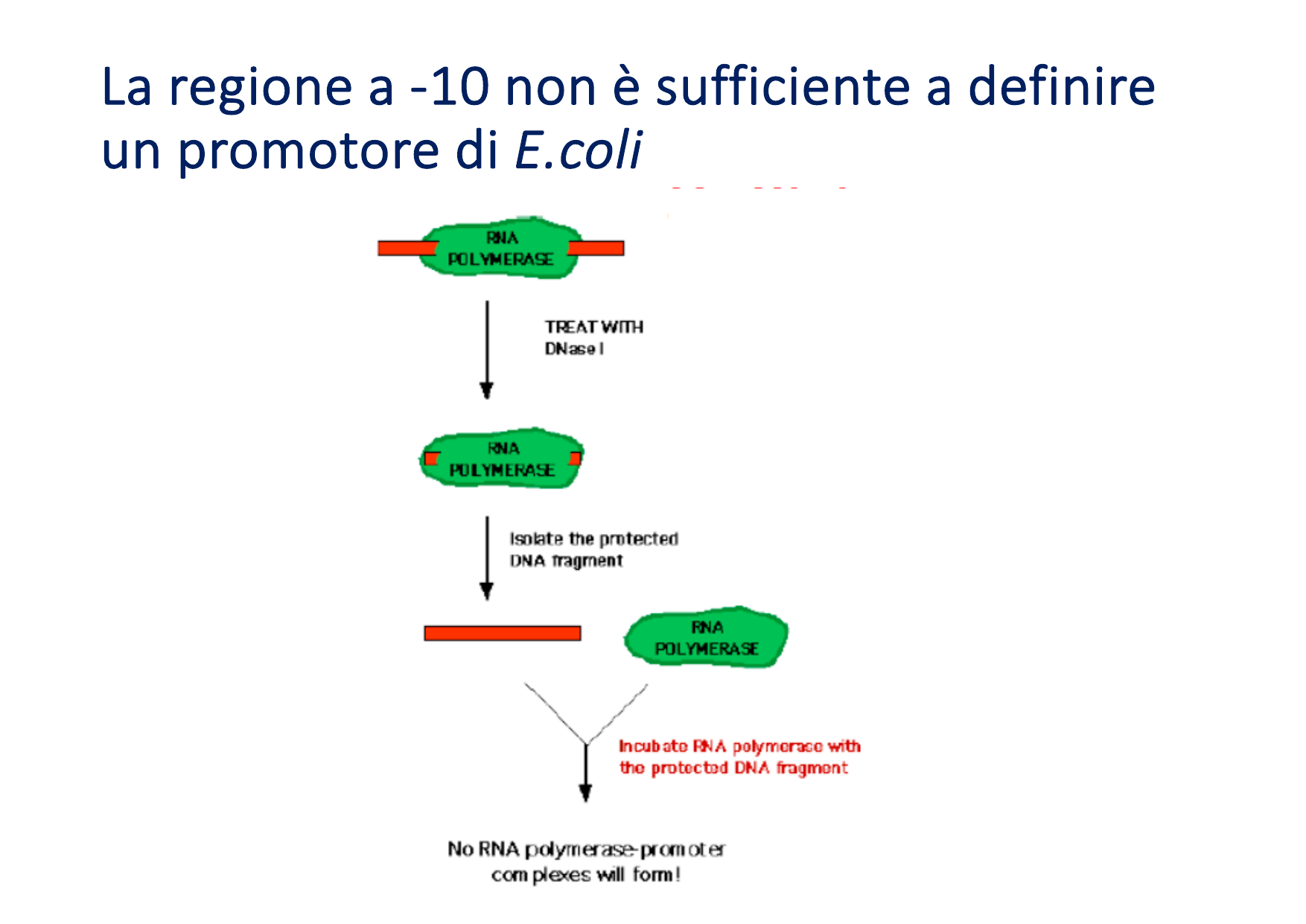
- Funzione della regione -35 (Sito di Aggancio): Se mutata, l’RNA polimerasi non riesce a formare il complesso chiuso. È la sequenza fondamentale per il primo riconoscimento e per la stabilità del legame iniziale dell’enzima al DNA.
- Funzione della regione -10 (Sito di Denaturazione): Se mutata, l’RNA polimerasi si lega ma non riesce a denaturare il doppio filamento per formare il complesso aperto (la bolla di trascrizione). Una presenza eccessiva di Citosine e Guanine (C-G) in questa regione aumenta la stabilità termodinamica del DNA, rendendo più difficile l’apertura e riducendo drasticamente l’efficienza trascrizionale.
Forza del Promotore e Regolazione
La forza di un promotore indica il numero di trascritti che vengono avviati da quella sequenza nell’unità di tempo.

Dipende da tre fattori termodinamici e cinetici principali:
- L’affinità di legame dell’RNA polimerasi per il promotore (maggiore quanto più le sequenze -10 e -35 sono vicine alla sequenza consenso).
- L’efficacia della transizione da complesso chiuso a complesso aperto (denaturazione locale).
- L’efficacia dell’allontanamento dell’enzima dal promotore dopo l’avvio della sintesi (promoter clearance).
Esempi di Forza e Regolazione
- Promotori deboli: L’operone LAC possiede un promotore naturalmente debole, lontano dalla sequenza consenso perfetta. Mutazioni spontanee che avvicinano la sua sequenza a quella consenso ne aumentano significativamente la forza.
- Elementi UP: Nei geni deputati alla sintesi di rRNA, dove è necessaria una trascrizione massiva, la cellula batterica presenta elementi regolatori aggiuntivi chiamati “elementi UP” a monte della regione -35 per incrementare ulteriormente l’affinità della polimerasi.
- Trascrizione Basale vs Regolata:
- Trascrizione basale: Mediata esclusivamente dall’interazione diretta tra la sequenza del promotore e l’oloenzima dell’RNA polimerasi.
- Trascrizione regolata: Richiede l’intervento di fattori di trascrizione aggiuntivi (attivatori o repressori) per modulare positivamente o negativamente l’attività dell’RNA polimerasi.
Anatomia del Promotore Eucariotico (Promotore di Classe 2)
Negli eucarioti, i promotori per l’RNA-polimerasi 2 (promotori di classe 2) sono localizzati a monte della porzione codificante del gene e si presentano strutturalmente suddivisi in aree distinte:
1. La Porzione Centrale (Core Promoter)
Il core promoter è la regione minima essenziale per l’assemblaggio del complesso di pre-inizio e l’avvio della trascrizione. È una sequenza breve situata a cavallo del sito di inizio trascrizionale, tipicamente compresa tra -50 e +50 paia di basi. Le sue funzioni principali sono:
- Permettere l’assemblaggio del complesso di pre-inizio.
- Stabilire con precisione il sito d’inizio della trascrizione (+1) e la direzione di trascrizione (definendo qual è il filamento stampo da leggere).
- Rispondere alle influenze regolatorie di attivatori o repressori legati ad altre regioni.
A differenza del promotore batterico (che richiede obbligatoriamente entrambe le sequenze consenso -10 e -35), nel core promoter eucariotico le sequenze consenso possono essere presenti singolarmente o in combinazioni variabili. È sufficiente una sola di queste sequenze per consentire l’avvio della trascrizione. Le sequenze principali sono:
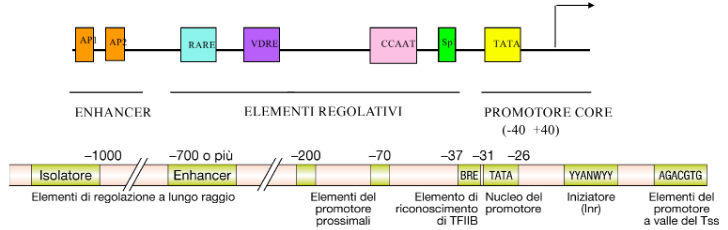
- TATA box: Caratterizzata dalla sequenza consensus TATAAA, situata tra le posizioni -31 e -26. (Nota: è l’elemento più rilevante e studiato del core promoter).
- Initiator (Inr): Situata a cavallo del sito di inizio, tra -2 e +4. È un promotore debole se presente da solo, e solitamente richiede l’ausilio di altre sequenze.
- DPE (Downstream Promoter Element): Localizzato a valle del sito d’inizio (già lungo il corso della trascrizione).
- BRE (TFIIB Recognition Element): Posizionato a monte rispetto alla TATA box.
Esperimento in Vitro sulla “Forza” delle Sequenze del Core Promoter
La funzionalità e la forza relativa delle sequenze del core promoter sono state analizzate in vitro inserendo in provetta il gene bersaglio, l’RNA polimerasi II, i fattori generali della trascrizione e i nucleotidi:
- Alterando la normale distanza in nucleotidi tra la TATA box e l’Initiator (spostando l’una o l’altro in avanti o indietro), si è osservato che il sito d’inizio della trascrizione (+1) si sposta seguendo la TATA box.
- Sperimentalmente, la TATA box si è dimostrata l’elemento consensus più forte del core promoter nel determinare la posizione d’inizio.
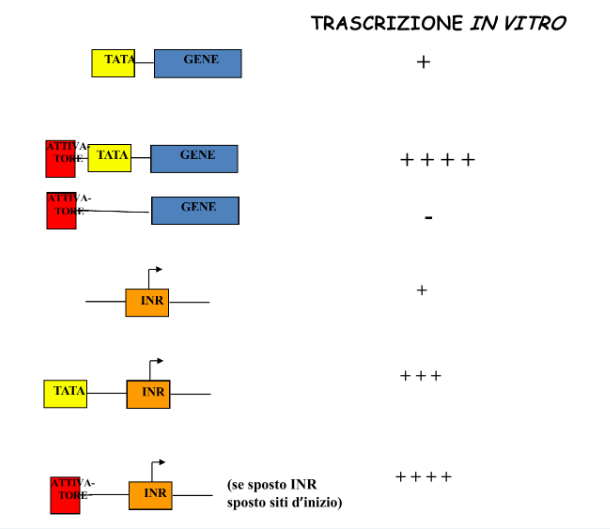

Nota: Sebbene in vitro il solo core promoter consenta una trascrizione basale minima, in vivo esso è inattivo a causa della struttura della cromatina altamente condensata (inibizione da nucleosomi). Per consentire la trascrizione sono necessari ulteriori elementi regolativi adiacenti che richiamino fattori capaci di decondensare la cromatina.
2. Le Porzioni Regolative
-
Promotore Prossimale (Regione Regolativa): Si estende a monte del core promoter fino a circa -400 paia di basi. Contiene elementi di sequenza che legano fattori di trascrizione regolativi, distinguibili in:
- Costitutivi: Sempre presenti nella cellula, facilitano costantemente l’avvio della trascrizione.
- Non costitutivi: Attivati solo in risposta a specifici segnali cellulari (es. presenza di ormoni o vitamine).
- Cooperazione trascrizionale: Poiché la cromatina ha una forte capacità repressiva, un singolo attivatore non costitutivo è spesso insufficiente. Pertanto, i fattori non costitutivi agiscono cooperativamente con i fattori costitutivi per superare l’inibizione nucleosomica.
-
Elementi Distali (Enhancer e Silencer): Regioni regolatrici poste a distanze molto elevate (oltre -700 bp).
- Nota di approfondimento: Per una trattazione dettagliata delle proprietà di promiscuità degli enhancer, dei silenziatori e del ruolo essenziale degli isolatori (insulator) nella prevenzione di oncogenesi accidentale in terapia genica, si veda la nota dedicata Enhancer, Silencer e Isolatori.
(Sezione espansa con: sbobina 15)
🔗 Collegamenti
- Gene — 📋 fa parte di
- Trascrizione — 🔗 stesso meccanismo / stessa via
- RNA Polimerasi Batterica — 🔄 diagnosi differenziale / confronto
- Operone — 🔗 stesso meccanismo / stessa via
- RNA Polimerasi Eucariotiche — 🔗 stesso meccanismo / stessa via
- Ciclo di Trascrizione Eucariotica — 🔗 stesso meccanismo / stessa via
- Complesso di Pre-inizio — 🔗 stesso meccanismo / stessa via
- Coda CTD e Mediatore — 🔗 stesso meccanismo / stessa via
- Enhancer, Silencer e Isolatori — 📋 fa parte di