RNA Polimerasi Batterica
L’RNA polimerasi batterica è il complesso multimerico enzimatico responsabile della catalisi della trascrizione nei procarioti. È stata isolata per la prima volta nel 1969 dal batterio E. coli mediante tecniche cromatografiche.
Struttura dell’Enzima
L’enzima si presenta in due conformazioni strutturali principali con funzioni distinte:
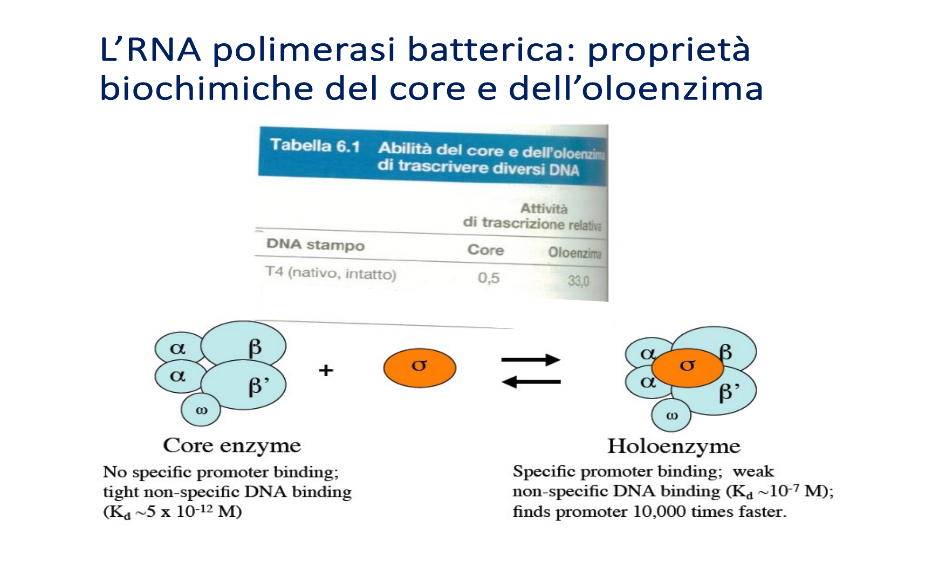
1. Core Enzimatico (Nucleo Centrale)
- Composizione: È composto da cinque subunità stabili: due subunità , una subunità , una subunità e una subunità .
- Subunità (violetto) e (blu): Rappresentano le “chele” dell’enzima che abbracciano il DNA stampo, formando un solco di alloggiamento. Dal punto di vista elettrostatico, sono caratterizzate da amminoacidi carichi positivamente per interagire con i gruppi fosfato (carichi negativamente) del DNA.
- Le due subunità : Hanno una funzione prevalentemente strutturale, servendo a tenere assemblato l’intero complesso.
- Subunità : Non svolge funzioni catalitiche o strutturali fondamentali per il processo trascrizionale.
- Proprietà biologiche: Presenta un’elevata affinità per le sequenze aspecifiche del DNA e scarsa affinità per il promotore. Ha la capacità catalitica di formare i legami fosfodiesterici e sintetizzare l’RNA, ma non è in grado di riconoscere dove iniziare la trascrizione. È la forma attiva nella fase di allungamento.
(Sezione espansa con: sbobina 11)
2. Oloenzima
- Composizione: È costituito dal core enzimatico associato a un fattore di specificità, la subunità (fattore sigma) (nota: la sbobina 10 menziona al rigo 68 il legame con una subunità , ma chiarisce successivamente al rigo 78 che sono le subunità ad associarsi al core libero).
- Proprietà biologiche: Has un’eccellente affinità per le sequenze del promotore (tramite legami deboli) e scarsissima affinità per il resto del DNA. Non trascrive aspecificamente. È la forma necessaria per la fase di inizio.
Canali dell’RNA Polimerasi
La struttura tridimensionale dell’RNA polimerasi definisce molteplici canali funzionali che regolano il transito dei substrati e dei prodotti:
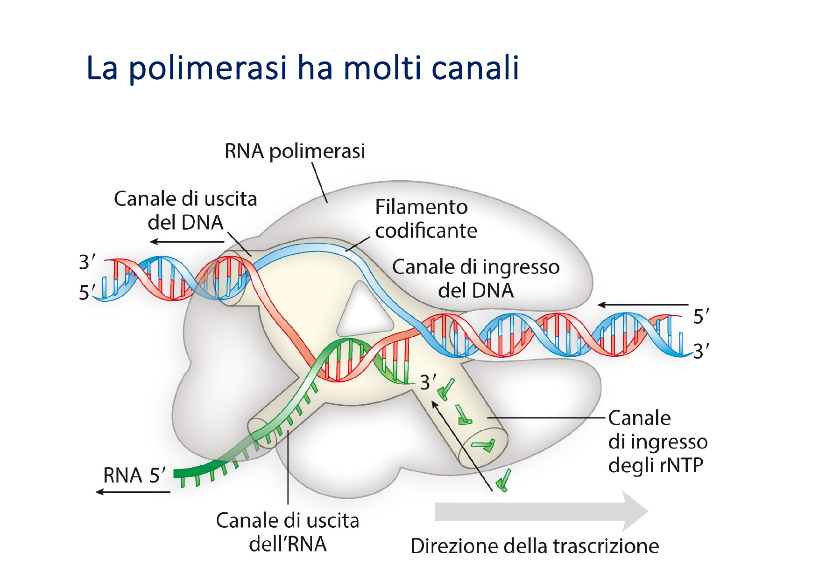
- Canale di ingresso del DNA: Corrisponde alla “chela” dell’enzima. Al suo interno, il DNA a doppio filamento incontra una struttura proteica chiamata incudine, che facilita e mantiene la denaturazione dei filamenti di DNA, garantendo l’esposizione del filamento stampo per l’appaiamento con i ribonucleotidi.
- Canale di ingresso degli rNTP: Permette l’accesso dei ribonucleotidi trifosfato liberi all’interno del sito attivo. Gli rNTP entrano in modo casuale e vengono incorporati solo se complementari alla base esposta sul DNA stampo. L’incorporazione è regolata dall’affinità chimica e dal tempo di permanenza del nucleotide nel sito attivo (l’enzima non “legge” attivamente, ma catalizza solo l’unione per complementarietà).
- Canale di uscita dell’RNA: Permette la fuoriuscita del filamento singolo di RNA neosintetizzato.
- Canale di uscita del DNA rinaturato: Consente l’uscita del doppio filamento di DNA che si richiude spontaneamente alle spalle della bolla di trascrizione.
(Sezione creata con: sbobina 11)
Fedeltà di Trascrizione e Sistemi di Correzione
L’RNA polimerasi possiede sistemi intrinseci di correzione degli errori di appaiamento:
- Frequenza di errore: La trascrizione presenta un tasso di errore superiore (minore accuratezza) rispetto alla replicazione operata dalla DNA polimerasi.
- Significato biologico: Questo divario di fedeltà è tollerato dalla cellula poiché:
- I trascritti di RNA hanno una vita media limitata (sono transitori e non permanenti).
- Un eventuale errore colpisce una singola molecola di RNA tra i numerosi trascritti corretti dello stesso gene, minimizzando l’impatto funzionale.
- Le mutazioni del DNA nella replicazione verrebbero invece trasmesse permanentemente a tutte le cellule figlie.
(Sezione creata con: sbobina 11)
Studi Sperimentali in Vitro
La scoperta delle proprietà funzionali del core e dell’oloenzima deriva da esperimenti di trascrizione in vitro combinando diverse forme dell’enzima e diversi stampi di DNA:
| Enzima | Stampo di DNA | Esito della Trascrizione |
|---|---|---|
| Core | DNA Umano (senza promotori batterici) | Trascrizione aspecifica e continua (punti di inizio casuali) |
| Oloenzima | DNA Umano | Nessuna trascrizione (l’oloenzima non si lega a sequenze aspecifiche) |
| Core | DNA di E. coli (con promotori) | Trascrizione casuale e discontinua |
| Oloenzima | DNA di E. coli | Trascrizione corretta e specifica (avviata esclusivamente dai promotori) |
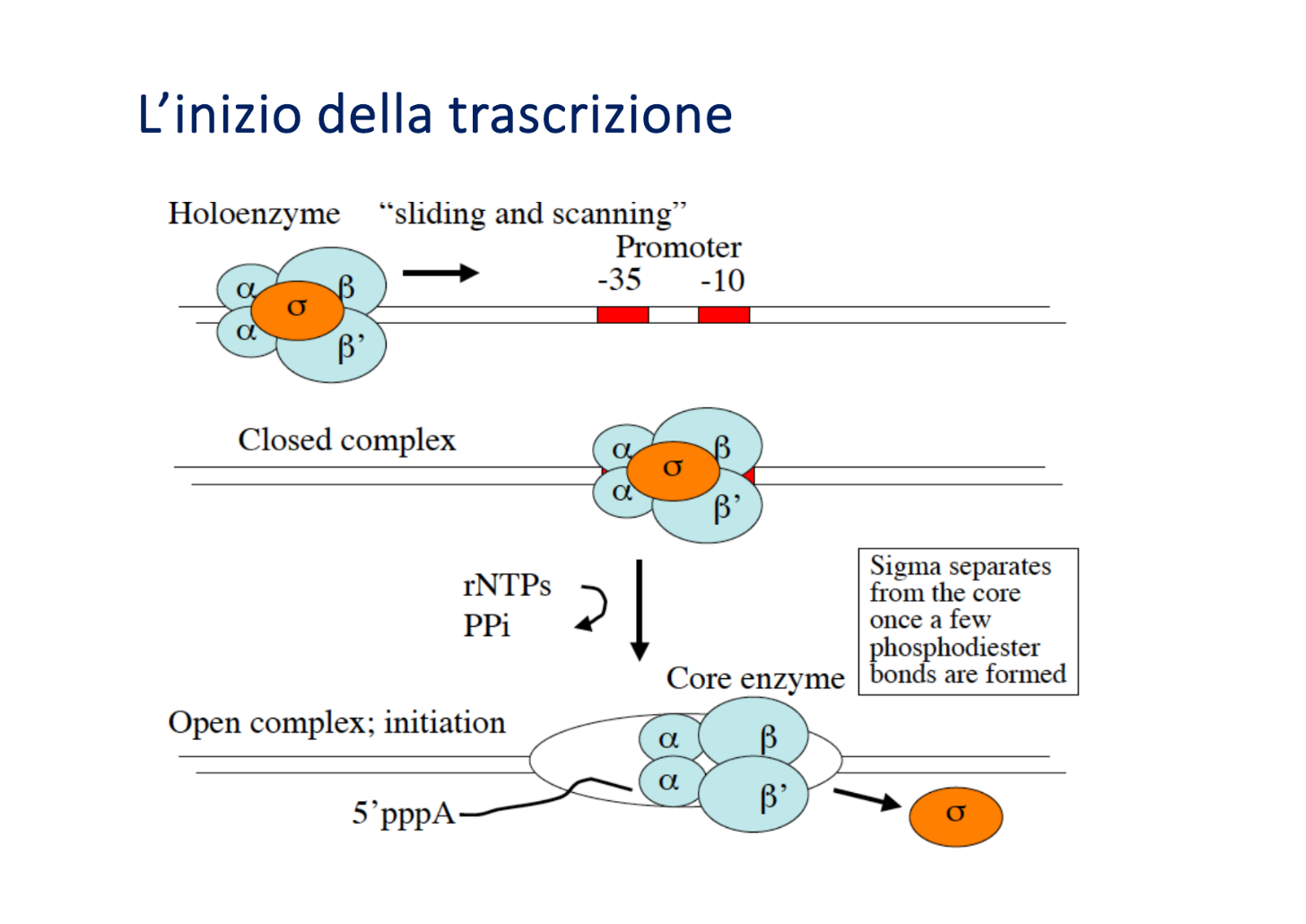
Meccanismo d’Azione e Ciclo del Fattore Sigma
La trascrizione procariotica segue un andamento ciclico basato sul passaggio tra oloenzima e core:
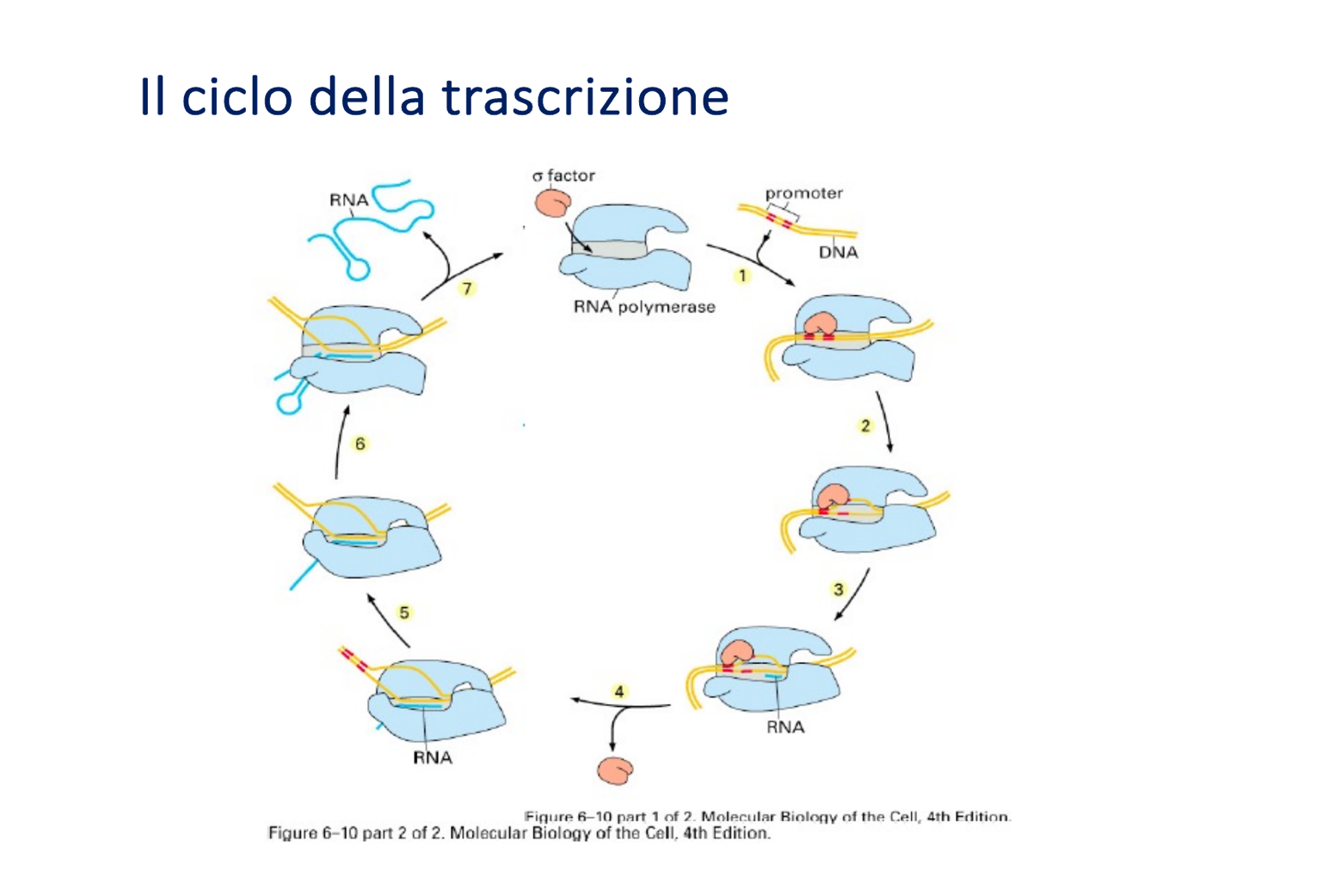
- Sliding and Scanning (Scansione): Nel citosol del batterio, le subunità libere si associano ai core enzimatici per formare l’oloenzima. Questo complesso si aggancia al DNA e lo “scansiona” muovendosi lungo il filamento (tramite scorrimento continuo o legami/distacchi successivi) finché non individua il promotore.
- Aggancio Stabile: Una volta riconosciuto il promotore, l’oloenzima vi si lega saldamente arrestandosi e formando il complesso chiuso.
- Denaturazione: L’enzima provoca l’apertura del doppio filamento (complesso aperto) senza l’intervento di elicasi esterne.
- Rilascio del Promotore: L’oloenzima sintetizza lentamente i primi nucleotidi (circa 12 legami fosfodiesterici). Poiché l’oloenzima ha un’altissima affinità per il promotore, l’allontanamento da esso richiede il distacco e rilascio della subunità .
- Allungamento: Una volta liberata la subunità , l’enzima torna nella conformazione di core, assume una forma chiusa (a “chela di granchio”) e procede rapidamente e in modo processivo lungo il DNA stampo fino al segnale di terminazione. Il core viene poi rilasciato nel citosol ed è pronto ad associarsi a una nuova subunità per ricominciare il ciclo.
🔗 Collegamenti
- Trascrizione — 📋 fa parte di
- Gene — 🔬 reperto diagnostico
- Promotore — 🔗 stesso meccanismo / stessa via
- Cellula Procariotica — 🔗 stesso meccanismo / stessa via