Proteina p53
La proteina p53 (codificata dal gene TP53) è il principale oncosoppressore nucleare, definito il “guardiano del genoma”. Rappresenta il gene più frequentemente mutato nelle forme tumorali dell’uomo. Svolge un ruolo cruciale nella coordinazione della risposta cellulare a diversi tipi di stress fisiologico o genotossico.
Struttura
p53 è una proteina organizzata in domini funzionali distinti; conoscere la struttura chiarisce perché certe mutazioni la inattivano (vedi sezione finale):
- Dominio di transattivazione (N-terminale): recluta la macchina trascrizionale ed è il sito su cui MDM2 si lega per inibirla.
- Dominio di legame al DNA (DBD, centrale): riconosce le sequenze specifiche dei geni bersaglio. È la regione dove cadono la maggior parte delle mutazioni puntiformi inattivanti nei tumori umani.
- Dominio di oligomerizzazione (C-terminale): consente l’assemblaggio di quattro monomeri nel tetramero, la forma funzionalmente attiva come fattore di trascrizione.
- Segnale di localizzazione nucleare (NLS): indispensabile per l’import nel nucleo, dove p53 svolge la sua funzione.
p53 funziona quindi come tetramero: serve sia il corretto legame al DNA (DBD) sia la tetramerizzazione (C-terminale) sia l’ingresso nel nucleo (NLS). La perdita di uno qualsiasi di questi requisiti la rende inefficace.
Segnali Attivatori (Stress Cellulare)
L’attivazione di p53 è indotta da molteplici fattori intrinseci ed estrinseci:
- Danni al DNA e stress genotossico: Rotture del doppio filamento di DNA.
- Stress replicativo: Anomalie o blocchi nella duplicazione del DNA.
- Disfunzione del fuso mitotico: Anomalie nella segregazione cromosomica.
- Erosione dei telomeri: Raggiungimento della senescenza replicativa.
- Attivazione aberrante di oncogeni: Espressione di oncogeni che stimolano eccessivamente il ciclo.
- Fattori estrinseci ambientali: Condizioni di ipossia (carenza di ossigeno).
- Segnali biochimici: Aumento delle concentrazioni di ossido nitrico (NO) o delezione del pool di ribonucleotidi liberi intracellulari.
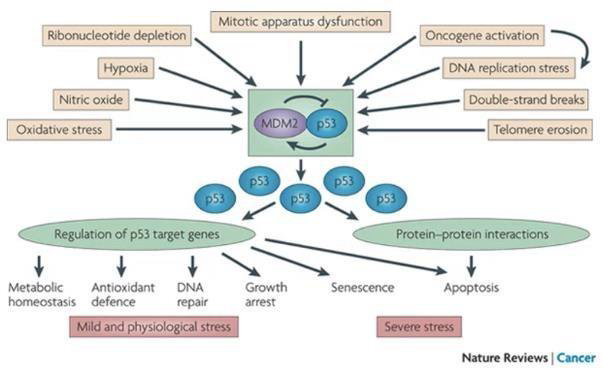
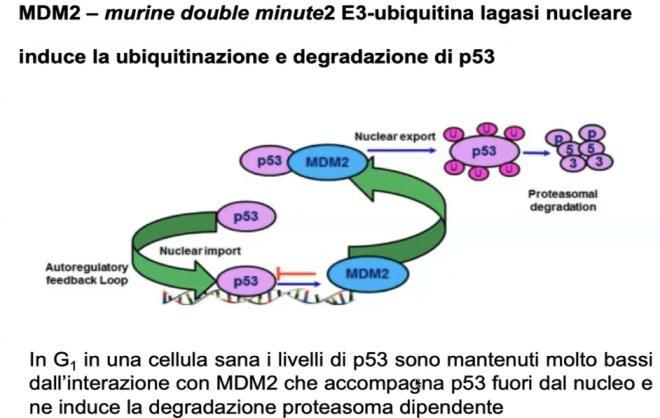
Regolazione Fisiologica: Il Loop p53-MDM2
In condizioni fisiologiche normali (assenza di stress), l’attività di p53 deve essere mantenuta estremamente bassa per evitare l’arresto del ciclo o l’apoptosi spontanea:
- Associazione con MDM2: Nel citoplasma, p53 si associa al suo inibitore principale MDM2 (murine double minute-2), una proteina avente attività enzimatica di ubiquitina ligasi E3.
- Cosa significa “E3”: l’ubiquitinazione richiede una cascata di tre enzimi — E1 (attivante, carica l’ubiquitina consumando ATP), E2 (coniugante, la trasporta) ed E3 (ligasi, l’ultimo della catena). È l’E3 a riconoscere il substrato specifico e a trasferirvi l’ubiquitina: è quindi l’enzima che dà specificità al sistema. MDM2 è l’E3 dedicata a p53. (Per il meccanismo generale ubiquitina–proteasoma vedi Modificazioni Post-Traduzionali.)
- Poli-ubiquitinazione e Degradazione: MDM2 lega p53 e vi catalizza l’aggiunta di una catena di ubiquitina (poli-ubiquitinazione).
- Destino della Proteina: La poli-ubiquitinazione agisce come segnale che impedisce l’accumulo nucleare di p53 (promuovendone l’esportazione dal nucleo o la ritenzione citoplasmatica) e ne determina la rapida degradazione nel proteasoma. Ciò assicura che i livelli di trascrizione dei geni target di p53 rimangano basali.
Meccanismo di Attivazione e Risposta
Quando la cellula subisce uno stress:
- Fosforilazione di p53: Chinasi attivate dal danno (es. ATM/ATR in risposta a rotture del DNA) fosforilano p53.
- Distacco da MDM2: La fosforilazione altera la conformazione di p53, che perde la capacità di interagire con MDM2.
- Accumulo Nucleare: p53 non viene più ubiquitinata né degradata; la sua emivita aumenta drasticamente ed essa accumula nel nucleo in forma attiva.
Funzioni e Geni Target
Una volta accumulata nel nucleo, p53 agisce principalmente come fattore di trascrizione, modulando l’espressione di numerosi geni target coinvolti in:
- Arresto della crescita cellulare: Blocco del ciclo (es. inducendo il CKI p21) per consentire la riparazione.
- Riparazione del DNA: Attivazione degli enzimi riparatori delle lesioni.
- Induzione dell’apoptosi: Se il danno è irreparabile, p53 promuove la via intrinseca (regolando proteine della famiglia Bcl-2 e interagendo direttamente in complessi proteina-proteina pro-apoptotici).
- Senescenza: Blocco replicativo permanente.
- Inibizione dell’angiogenesi: Blocco dello sviluppo di nuovi vasi sanguigni.
- Regolazione del metabolismo energetico.
- Difesa dallo stress ossidativo.
Come p53 “decide”: riparabile vs irreparabile
La nota madre Sistemi di Controllo del Ciclo Cellulare descrive l’esito (“se irreparabile → apoptosi”) come una scatola nera. Il meccanismo con cui p53 distingue i due casi non è un singolo interruttore on/off, ma una dinamica temporale del segnale (codifica digitale, “a impulsi”):
- Il sensore a monte misura la durata del danno. La chinasi ATM si attiva in modo switch-like quando le proteine di riparazione si legano alle rotture del doppio filamento, e resta attiva finché il danno persiste. Più a lungo dura la riparazione, più a lungo ATM segnala.
- p53 risponde a pulsazioni, non a un livello costante. ATM induce impulsi ripetuti di p53 di ampiezza e periodo fissi (~5–6 ore ciascuno): è il numero di impulsi, non la loro intensità, a portare l’informazione sulla gravità del danno.
- Danno lieve/riparabile: pochi impulsi → la riparazione si completa, ATM si spegne, gli impulsi cessano e la cellula rientra nel ciclo.
- Danno grave/irreparabile: gli impulsi si susseguono e, superata una soglia critica, la dinamica passa da pulsatile a un livello alto e sostenuto di p53 → apoptosi.
- Lo stesso p53 cambia “programma” di geni bersaglio. Nelle prime fasi p53 agisce da “arrester” inducendo p21 (arresto del ciclo + tempo per riparare); con il persistere del danno commuta verso un profilo “killer”, attivando geni pro-apoptotici (es. p53AIP1, regolatori della famiglia Bcl-2). Il ritardo evita un’apoptosi prematura prima che la riparazione abbia avuto una possibilità.
In sintesi: la gravità del danno è cronometrata (durata dell’attività ATM), tradotta nel conteggio degli impulsi di p53, e convertita in destino cellulare dal tipo di geni bersaglio che p53 accende.
Meccanismi Mutazionali di Perdita di p53
La perdita o l’inattivazione della funzione di p53 nelle cellule neoplastiche può avvenire attraverso diverse modalità:
-
Mutazioni puntiformi nel dominio di legame al DNA (DBD): La sostituzione di singoli amminoacidi nei domini responsabili del legame con le sequenze specifiche del DNA impedisce a p53 di agire come fattore di trascrizione sui suoi geni target. Questa è la causa principale di inattivazione riscontrata nei tumori umani.
(Integrazione da: sbobina 27)
- Mutazioni Hot-Spot C-G T-A: Quasi il 50% di queste mutazioni inattivanti a carico del gene TP53 sono transizioni C T indotte dalla deaminazione spontanea di residui di 5-metilcitosina (vedi Metilazione del DNA).
-
Perdita della porzione carbossi-terminale (C-terminale): Rimuove il dominio di oligomerizzazione della proteina, inibendo la formazione del tetramero p53 funzionale.
-
Sovraespressione di MDM2: L’amplificazione del gene MDM2 causa un aumento patologico dell’ubiquitinazione e della degradazione di p53, portando a perdita funzionale anche in assenza di mutazioni su TP53 (tipico di alcuni tumori del SNC e di diversi sarcomi).
-
Infezioni virali: Proteine prodotte da virus oncogeni (come l’HPV nella cervice uterina, l’HBV nel fegato e l’EBV in alcuni linfomi) si legano a p53 promuovendone la degradazione o sequestrandola.
-
Blocco dell’import nucleare: La proteina p53 perde la capacità di migrare nel nucleo e rimane sequestrata nel citoplasma, risultando inefficace come fattore trascrizionale (osservato in neuroblastomi e carcinomi mammari).
Fonti
- Struttura, loop p53-MDM2, geni target e meccanismi mutazionali: sbobine del corso (16, 27).
- Dinamica a impulsi e decisione riparabile/irreparabile (codifica digitale, p53 “arrester”/“killer”): letteratura — Cell fate decision mediated by p53 pulses (PNAS, PMC2710988); Two-phase dynamics of p53 in the DNA damage response (PNAS, PMC3107314).
🔗 Collegamenti
- Oncosoppressori — 📋 fa parte di
- Apoptosi — ⬆️ causa
- Via Intrinseca dell’Apoptosi — 🔗 stesso meccanismo
- Sistemi di Controllo del Ciclo Cellulare — 🔗 stesso meccanismo
- Modificazioni Post-Traduzionali — 🔄 connessione molecolare (ubiquitinazione)
- Metilazione del DNA — ⬆️ causa