Metilazione del DNA
La metilazione del DNA è una modificazione chimica post-replicativa del genoma che consiste nell’aggiunta covalente di un gruppo metilico alle basi azotate. Rappresenta il primo meccanismo epigenetico caratterizzato storicamente ed è fondamentale per la regolazione dell’espressione genica e il mantenimento della stabilità genomica.
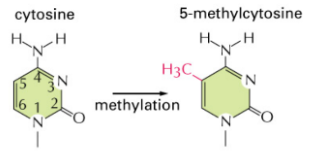
Ad oggi, nel DNA vengono riconosciute sei basi, tra cui la 5-metilcitosina e la 5-idrossimetilcitosina, che derivano dalla modificazione chimica della citosina.
Meccanismo Chimico ed Enzimi
La metilazione avviene in seguito all’incorporazione dei nucleotidi standard da parte della DNA polimerasi (non vengono incorporati nucleotidi pre-metilati).
- Enzimi Scrittori: La modificazione è catalizzata dalle DNA metiltransferasi (DNMT), enzimi che trasferiscono un gruppo metilico da un donatore chimico specifico alla citosina.
- Specificità di Base:
- Nei mammiferi, la metilazione avviene esclusivamente a carico delle citosine (nello specifico sul carbonio in posizione 5, generando la 5-metilcitosina).
- Nei procarioti, la metilazione può avvenire sia sulla citosina che sull’adenina.
Siti di Metilazione: CG vs CH
Storicamente si riteneva che la metilazione avvenisse solo a livello del dinucleotide CG (una citosina seguita da una guanina in direzione 5’ 3’). Oggi si distinguono due tipologie:
- Metilazione CG: È di gran lunga l’evento di metilazione più comune e diffuso nelle cellule somatiche (coinvolge dal 60% al 90% di tutti i dinucleotidi CG del genoma).
- Metilazione non-CG (o Metilazione CH): In cui la citosina è seguita da un nucleotide diverso dalla guanina (H = A, T o C). Questo pattern è caratteristico delle cellule staminali embrionali (ESC) e dei neuroni, risultando particolarmente abbondante nel cervello adulto per potenziare la complessità regolatoria dell’espressione genica.
Funzioni Biologiche della Metilazione
La metilazione del DNA svolge ruoli cruciali nello sviluppo e nella fisiologia cellulare:
-
Regolazione Trascrizionale (Silenziamento): La metilazione a livello dei promotori genici correla con il silenziamento della trascrizione (promotore metilato = gene spento; promotore non metilato = gene espresso). Questo meccanismo riguarda i geni regolati tessuto-specifici, mentre i geni costitutivi (housekeeping) presentano promotori costantemente non metilati.
Meccanismi di Repressione Trascrizionale
La metilazione può reprimere la trascrizione attraverso due modelli principali:
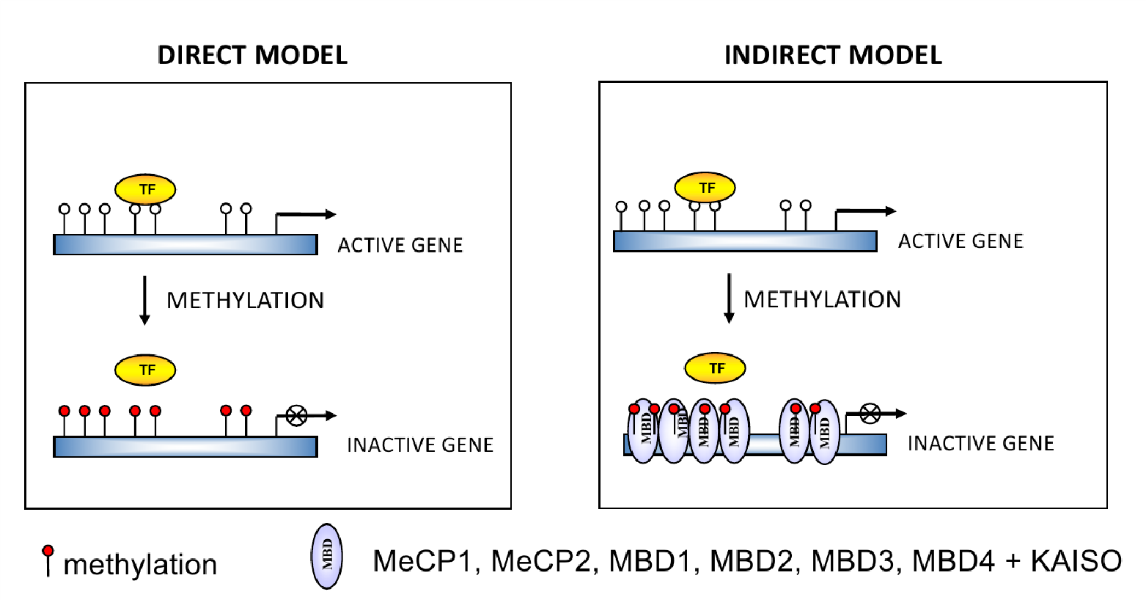
- Modello Diretto (Ingombro Sterico): Il gruppo metilico della 5-metilcitosina sporge direttamente nel solco maggiore del DNA. Poiché la maggior parte dei fattori di trascrizione inserisce la propria alfa-elica di legame nel solco maggiore, la presenza del gruppo metilico impedisce fisicamente il legame del fattore alla sua sequenza di riconoscimento. Questo meccanismo presenta dei limiti biologici:
- Alcuni fattori riescono a legare la sequenza anche quando metilata (sebbene il gene resti comunque represso).
- La sequenza bersaglio dei fattori di trascrizione è tipicamente di 10-12 paia di basi, mentre il dinucleotide CG si trova statisticamente solo una volta ogni 100 dinucleotidi, rendendo molto bassa la probabilità che un dinucleotide CG si trovi esattamente nel sito di legame specifico.
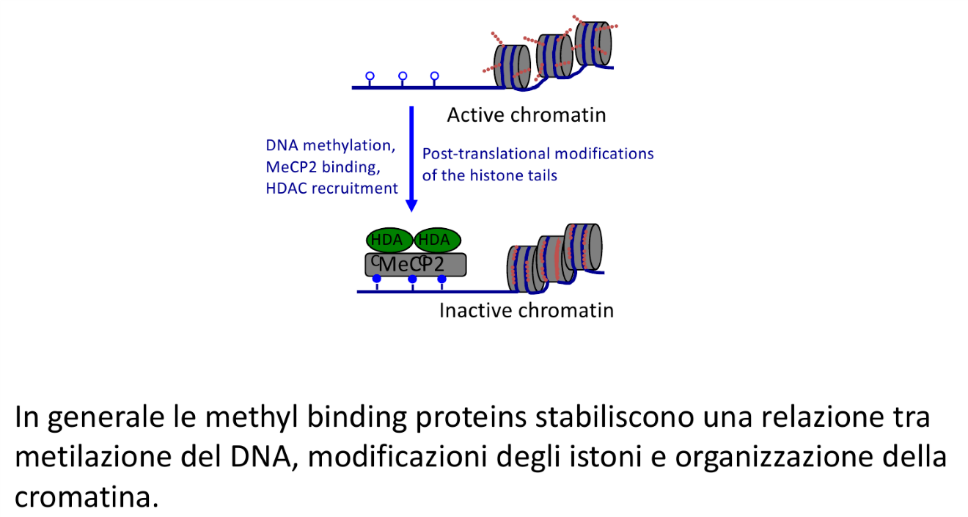
- Modello Indiretto (Reclutamento di Lettori): Proposto da Adrian Bird per spiegare come promotori sprovvisti di dinucleotidi CG nel sito specifico di legame vengano comunque repressi. In questo modello, i gruppi metilici attirano proteine “lettrici” specifiche definite Proteine MBD (o methyl binding proteins), come Proteina MeCP2. Queste proteine si legano al DNA metilato e inibiscono la trascrizione tramite un meccanismo attivo di repressione, richiamando modificatori istonici che inducono la chiusura della cromatina (eterocromatina).
- Modello Diretto (Ingombro Sterico): Il gruppo metilico della 5-metilcitosina sporge direttamente nel solco maggiore del DNA. Poiché la maggior parte dei fattori di trascrizione inserisce la propria alfa-elica di legame nel solco maggiore, la presenza del gruppo metilico impedisce fisicamente il legame del fattore alla sua sequenza di riconoscimento. Questo meccanismo presenta dei limiti biologici:
-
Difesa Genomica e Stabilità: La maggior parte della metilazione genomica si concentra su sequenze altamente ripetute e parassitarie (trasposoni, retrovirus, sequenze LINE e SINE). Il silenziamento tramite metilazione impedisce la trascrizione e la trasposizione di questi elementi mobili, prevenendo la mutagenesi da inserzione.
- Confronto evolutivo: Il moscerino della frutta (Drosophila melanogaster) ha perso quasi totalmente la metilazione del DNA e, di conseguenza, soffre di un’elevata frequenza di mutagenesi indotta dal salto dei trasposoni. L’essere umano, pur possedendo un carico enorme di sequenze trasponibili, non soffre di mutagenesi da trasposizione attiva grazie al silenziamento operato dalla metilazione.
(Integrazione da: sbobina 28)
- In cellule prive della DNA metiltransferasi principale (DNMT1), si osserva un incremento dell’attività di trasposizione.
- Nel cervello di soggetti affetti da Sindrome di Rett (privi del lettore MeCP2 funzionante), la cromatina risulta meno compatta e si riscontra un aumento anomalo della mobilità dei trasposoni, confermando l’importanza di questa difesa genomica.

-
Inattivazione del Cromosoma X: Necessaria per il dosaggio genico nelle cellule femminili.
-
Controllo dell’Imprinting Genico: Regolazione allele-specifica in base all’origine parentale.
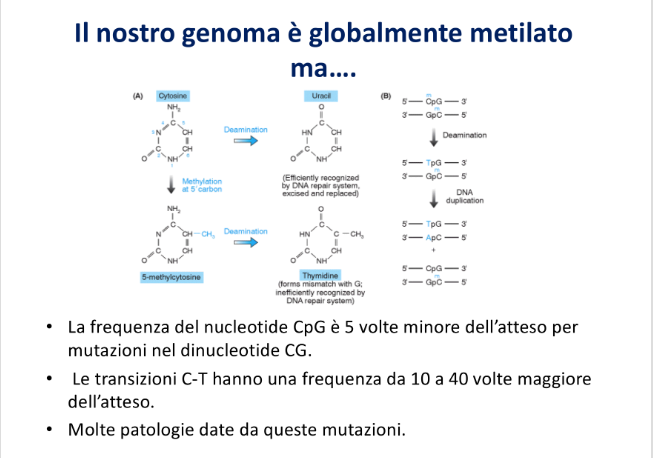
Deaminazione Spontanea della 5-Metilcitosina
La presenza di citosine metilate rappresenta un fattore di instabilità chimica che ha plasmato l’evoluzione del genoma:
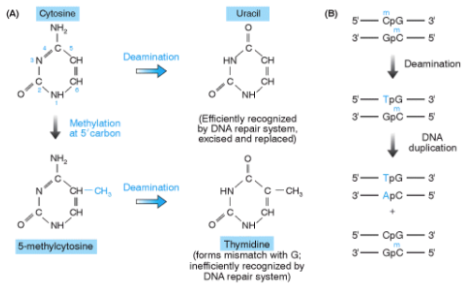
- Citosina non metilata: La sua deaminazione spontanea produce uracile. Poiché l’uracile non è una base fisiologica del DNA, viene prontamente riconosciuto e rimosso dai sistemi di riparazione cellulari senza conseguenze mutazionali.
- 5-Metilcitosina (Citosina metilata): La sua deaminazione spontanea rimuove un gruppo amminico trasformandola in timina (timidina). Poiché la timina è una base standard del DNA, questo evento genera un mismatch T-G subdolo e difficile da rilevare per la cellula.
Scenari del Mismatch T-G
- Scenario 1 (Riparazione): I sistemi di riparazione riconoscono l’appaiamento anomalo T-G, rimuovono la timina errata sul filamento e la sostituiscono con una citosina, ripristinando la coppia C-G originale.
- Scenario 2 (Fissazione della mutazione): Se la cellula si replica prima che avvenga la riparazione, i due filamenti si separano. Il filamento non mutato genera una molecola figlia corretta (C-G), mentre il filamento con la timina mutata incorpora un’adenina di fronte ad essa, fissando permanentemente una transizione C-G T-A ereditabile.
Conseguenze Evolutive e Patologiche
- Depauperamento dei dinucleotidi CG: A causa di questo meccanismo evolutivo, i dinucleotidi CG sono circa 5 volte meno frequenti di quanto atteso statisticamente (1 ogni 100 nucleotidi invece di 1 ogni 16).
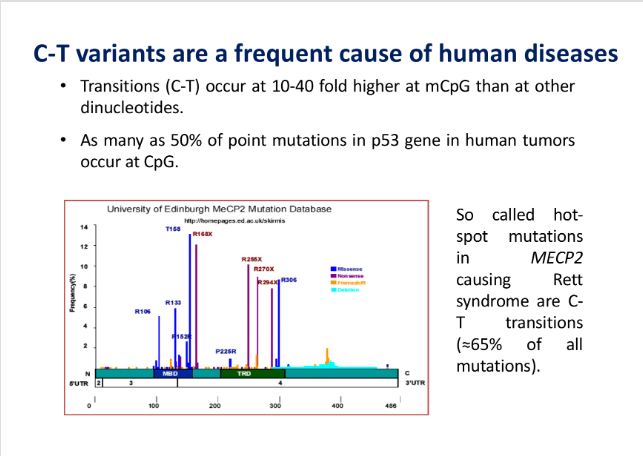
- Hot-spot Mutazionali: Le transizioni C T sono da 10 a 40 volte più frequenti a livello delle citosine metilate. Ad esempio, nel gene oncosoppressore p53, quasi il 50% delle mutazioni somatiche riscontrate nei tumori sono transizioni C T causate da questo meccanismo. Mutazioni analoghe inattivanti il gene MECP2 (o MCP2, lettore della metilazione) causano gravi patologie del neurosviluppo.
Evidenze Sperimentali d’Importanza (Adrian Bird)
L’assoluta necessità della metilazione per gli organismi multicellulari complessi è stata dimostrata mediante studi knockout e colture in vitro:
- Letalità In Vivo (Topo Knockout per DNMT1): La delezione del gene per la DNA metiltransferasi principale (DNMT1) in modelli murini determina l’arresto precoce dello sviluppo embrionale (subito dopo la gastrulazione) e la morte dell’embrione in utero.
- Proliferazione In Vitro (Cellule Staminali Embrionali): Le cellule staminali embrionali (ESC) isolate da embrioni privi di DNMT1 sono in grado di sopravvivere e proliferare normalmente in vitro. L’analisi del loro genoma evidenzia la perdita del 95% della metilazione complessiva, lasciando un 5% di metilazione residua che dimostra l’esistenza di altre DNA metiltransferasi (de novo).
- Morte al Differenziamento In Vitro: Se si stimolano le ESC KO per DNMT1 a differenziarsi in vitro (es. in neuroni, epatociti, condrociti), le cellule non riescono a completare il programma di differenziamento e vanno incontro a morte cellulare programmata per apoptosi o necrosi.
- Differenze tra Specie:
- Le ESC di roditore (topo) sopravvivono e si dividono anche in assenza di tutte e tre le DNA metiltransferasi note (0% di metilazione).
- Le ESC umane non possono sopravvivere né proliferare in assenza di un livello minimo di metilazione del DNA, evidenziando una maggiore dipendenza regolatoria legata alla complessità della nostra specie.

5. Conservazione Evolutiva delle Metiltransferasi
(Integrazione da: sbobina 28) La metilazione del DNA è presente nei procarioti, assente nel lievito, scarsa in Drosophila ed abbondante nei vertebrati. Il sequenziamento genico ha mostrato che le DNA metiltransferasi umane sono omologhe a quelle di Escherichia coli. Questo dimostra che la metilazione è un meccanismo ancestrale antichissimo, mantenuto evolutivamente nei lignaggi in cui è rimasto utile (come i vertebrati per il controllo dello sviluppo) e perso secondariamente negli organismi (come lievito e drosofila) in cui non era altrettanto necessario.
Rappresentazione Grafica della Metilazione (Lollipop)
Negli studi genomici, lo stato di metilazione delle citosine in una determinata regione viene rappresentato graficamente tramite diagrammi a “lollipop” (stanghette verticali sormontate da un cerchio):
- Cerchio vuoto (bianco): Dinucleotide CG non metilato.
- Cerchio pieno (nero): Dinucleotide CG metilato.
- Cerchio diviso a metà: Stato di emimetilazione (metilato su un solo filamento) o metilazione allele-specifica (un allele metilato e l’altro no).
- Cerchio grigio: Metilazione parziale con frequenza intermedia nella popolazione cellulare.
Ereditarietà e Replicazione: Mantenimento vs De Novo
La replicazione del DNA è semiconservativa. Quando una doppia elica metilata simmetricamente viene duplicata, si generano due doppie eliche emimetilate (il filamento parentale conserva il gruppo metilico, il filamento di nuova sintesi ne è privo).
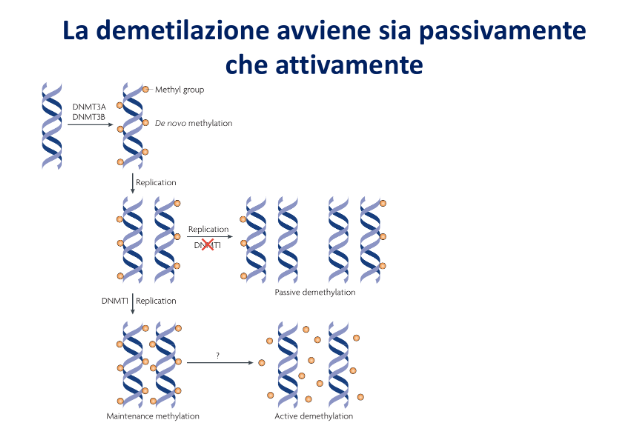
A questo punto intervengono due classi di attività metiltransferasica:
- Metilazione di Mantenimento: Operata principalmente da DNMT1. Riconosce specificamente il DNA emimetilato e aggiunge il gruppo metilico sul filamento di nuova sintesi di fronte alla citosina metilata parentale, ripristinando la simmetria del pattern originale nelle cellule figlie.
- Metilazione De Novo: Operata da altre DNMT. Aggiunge gruppi metilici su sequenze precedentemente non metilate su entrambi i filamenti, consentendo l’acquisizione di nuovi profili epigenetici durante il differenziamento cellulare.
Se il meccanismo di mantenimento viene inibito o non è attivo durante i cicli replicativi, il gruppo metilico viene progressivamente diluito ad ogni divisione cellulare in modo passivo (demetilazione passiva). In alternativa, la rimozione dei gruppi metilici può avvenire attivamente ad opera di enzimi demetilasi o “eraser” (demetilazione attiva), essenziale nei neuroni post-mitotici che devono rimodellare il proprio epigenoma pur non replicandosi più.
🔗 Collegamenti
- Codice Epigenetico — 📋 fa parte di
- Isole CpG — 🔄 diagnosi differenziale / confronto
- Dinamica della Metilazione del DNA nello Sviluppo — ➡️ evoluzione / progressione
- Mutazioni — ⬆️ causa
- Proteina p53 — ⬇️ conseguenza
- Cellule Staminali Pluripotenti Indotte — 🔄 connessione molecolare
- Apoptosi — ⬇️ conseguenza
- Necrosi — ⬇️ conseguenza
- Enzimi di Restrizione — 🔗 stesso meccanismo
- Proteine MBD — 🔗 stesso meccanismo
- Proteina MeCP2 — 🔗 stesso meccanismo
- Sindrome di Rett — ⬇️ conseguenza
- Sindrome ICF — ⬇️ conseguenza
- Metilazione del DNA nel Cancro — ➡️ evoluzione / progressione