Spliceosoma
Lo spliceosoma è il grande complesso macromolecolare deputato a catalizzare la rimozione degli introni dal pre-mRNA. Trattandosi di un processo in cui ogni singolo nucleotide è cruciale (un errore di sfasamento di una sola base compromette l’intera catena di lettura dell’mRNA), lo spliceosoma garantisce un’accuratezza estrema nel taglio e nella saldatura.
Struttura dello Spliceosoma
Lo spliceosoma è una ribonucleoproteina (RNP) costituita dall’interazione di proteine e piccoli RNA stabili:
- Subunità: È composto da sei subunità proteiche.
- snRNA (Small Nuclear RNA): Cinque di queste sei subunità contengono corti filamenti di RNA (lunghi circa un centinaio di nucleotidi) denominati piccoli RNA nucleari. Questi snRNA sono altamente conservati nel corso dell’evoluzione.
- snRNP: I complessi formati da proteine e snRNA prendono il nome di ribonucleoproteine nucleari (snRNP, comunemente chiamate “snurp”).
- Divisione del lavoro: Gli snRNA svolgono il ruolo catalitico e di riconoscimento di sequenza principale, mentre le proteine del complesso hanno una funzione strutturale e di facilitazione del corretto assemblaggio spaziale.
Meccanismo di Riconoscimento e Funzionamento
La straordinaria precisione dello spliceosoma nel riconoscere le sequenze consenso si basa sull’appaiamento complementare delle basi (legami idrogeno Watson-Crick) tra i suoi snRNA e il pre-mRNA da processare.
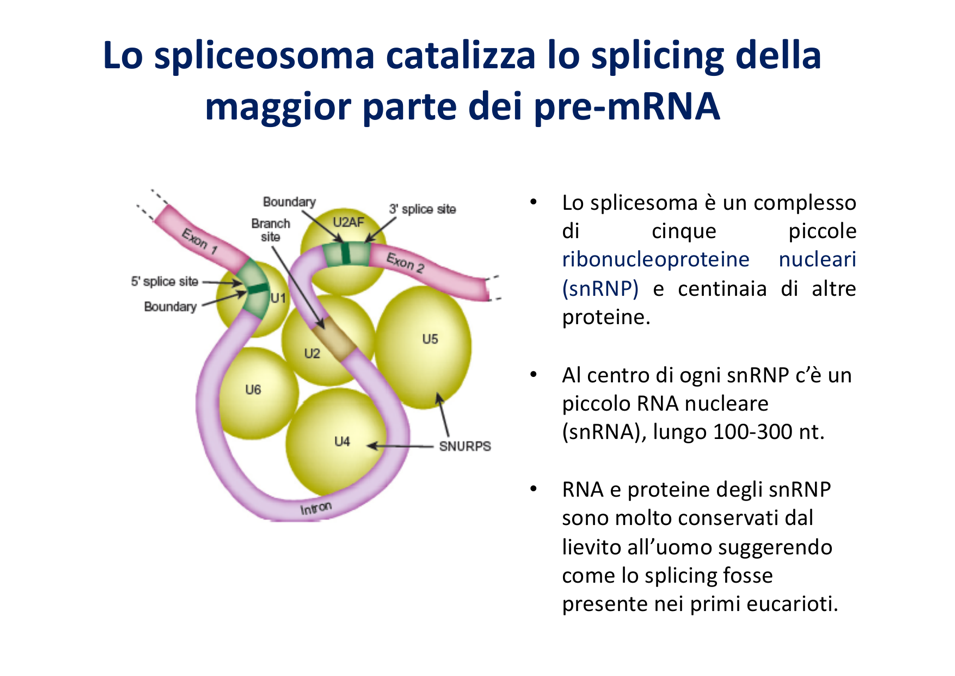
Assemblaggio ed Estroflessione dell’Introne
- Riconoscimento del 5’-Cap / Sito Donatore: La subunità contenente lo snRNA U1 si posiziona sul confine esone-introne al 5’ (sito donatore). L’estremità dello snRNA U1 è perfettamente complementare alla sequenza consensus GU, consentendo l’appaiamento.
- Riconoscimento del 3’ / Sito Accettore: La subunità proteica UBAF (biologicamente nota come U2AF) riconosce e si lega al sito accettore 3’ (AG).
- Piegamento e Connessione: Altre subunità dello spliceosoma si legano al branch point e fungono da ponte per connettere fisicamente la subunità al 5’ con quella al 3’.
- Catalisi: L’avvicinamento forzato dei due siti causa l’estroflessione dell’introne (che viene “spinto fuori” indipendentemente dalla sua lunghezza). Questo ripiegamento spaziale espone e attiva il gruppo ossidrile dell’adenina del branch point, avviando la reazione di transesterificazione (vedi Splicing).
Il Costo Energetico Nascosto
La catalisi chimica (le transesterificazioni) ha bilancio energetico neutro e non consuma ATP. L’ATP serve però all’assemblaggio: tra una tappa e l’altra lo spliceosoma rimodella il proprio nucleo, sostituendo e riposizionando gli snRNA. Questi riarrangiamenti sono operati da elicasi a RNA ATP-dipendenti (famiglie DEAD/DEAH-box), che sciolgono e riformano gli appaiamenti snRNA–pre-mRNA. Il complesso è quindi dinamico, non un assemblaggio statico.
Evidenze Sperimentali: Esperimenti di Recupero (Rescue)
Il ruolo degli snRNA nel riconoscimento specifico dei siti di splicing è stato confermato sperimentalmente dallo studio di mutazioni patogenetiche:
- Mutazione patogena: Una mutazione che altera la prima base conservata del sito donatore intronico (ad esempio, trasformando la sequenza consensus GU in AU) impedisce l’appaiamento di basi con lo snRNA U1 wild-type. Lo spliceosoma non riesce più a legarsi e lo splicing fallisce.
- Esperimento di Recupero (Rescue): Introducendo artificialmente in cellule in coltura uno snRNA U1 mutato modificato per essere complementare alla sequenza mutata del pre-mRNA (AU anziché GU), si osserva il ripristino del legame idrogeno e il recupero del corretto processo di splicing. Questa capacità competitiva dell’RNA modificato costituisce la base biologica diretta delle Terapie di Modulazione dello Splicing (snRNA U1 modificato e oligonucleotidi antisenso che competono con l’U1 fisiologico).
🔗 Collegamenti
- Splicing — 📋 fa parte di
- Capping e Poliadenilazione — 🔗 stesso meccanismo / stessa via
- Terapie di Modulazione dello Splicing — ➡️ evoluzione / progressione
- Elicasi — 🔗 stesso meccanismo / stessa via
- Difetti di Splicing — ⬇️ conseguenza