Splicing
Lo splicing dell’RNA è il processo di maturazione post-trascrizionale mediante il quale gli introni (sequenze non codificanti) vengono rimossi dal trascritto primario (pre-mRNA) e gli esoni (sequenze destinate al trascritto maturo) vengono saldati in sequenza per formare l’mRNA maturo. Questo meccanismo avviene esclusivamente nel nucleo delle cellule eucariotiche.
Scoperta dello Splicing ed Esperimenti Storici
La scoperta dello splicing (premiata con il Premio Nobel) ha svelato la struttura interrotta dei geni eucariotici, differenziandoli dai genomi compatti dei procarioti (es. E. coli), che sono quasi interamente codificanti e privi di splicing.
L’esperimento di Ibridazione e i Loop di DNA
La scoperta nacque dall’osservazione che l’RNA radiomarcato sintetizzato nel nucleo era quantitativamente molto superiore a quello che effettivamente raggiungeva il citoplasma. Per investigare questo fenomeno, i ricercatori:
- Denaturarono il DNA genomico umano a singolo filamento.
- Utilizzarono come sonda un trascritto di RNA maturo complementare a un gene noto (es. il gene della distrofina, ricco di introni).
- Ibridarono la sonda a RNA con il DNA genomico denaturato abbassando gradualmente la temperatura.
- All’osservazione al microscopio elettronico si notò che l’RNA si appaiava solo in corrispondenza di determinati tratti, mentre il DNA genomico formava vistose anse (loop) spaiate.
Questo dimostrò che:
- Le regioni appaiate corrispondevano agli esoni, conservati nel trascritto maturo.
- Le anse spaiate di DNA corrispondevano agli introni, sequenze presenti nel genoma ma assenti nell’mRNA maturo.
- La polimerasi trascrive l’intero gene (inclusi gli introni), che vengono poi rimossi progressivamente nel nucleo.
Sequenze Consensus dello Splicing
La specificità del taglio e della cucitura è garantita dal riconoscimento di precise sequenze segnale poste prevalentemente all’interno dell’introne e alle giunzioni esone-introne:
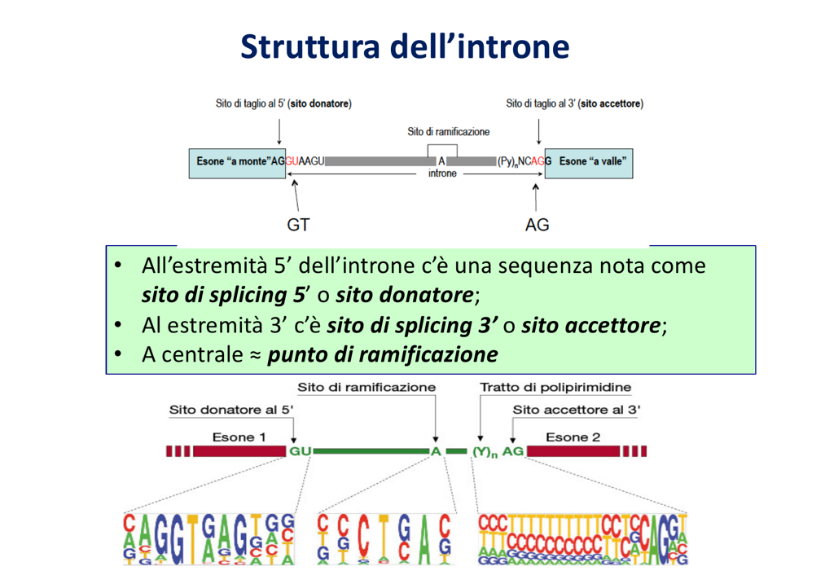
- Sito donatore al 5’: Localizzato all’estremità 5’ dell’introne (giunzione esone-introne), caratterizzato dalla sequenza dinucleotidica altamente conservata GU.
- Sito accettore al 3’: Localizzato all’estremità 3’ dell’introne (giunzione introne-esone), caratterizzato dalla sequenza dinucleotidica altamente conservata AG.
- Sito di ramificazione (Branch Point): Sequenza consensus situata all’interno dell’introne, contenente un residuo di adenina (A) altamente conservato fondamentale per l’avvio della reazione chimica.
Meccanismo Chimico: Le Transesterificazioni
Il processo di rimozione dell’introne avviene attraverso due reazioni chimiche consecutive di transesterificazione (scambio di legami fosfodiesterici tramite attacco nucleofilo sul fosforo), che non richiedono l’apporto di energia esterna (ATP):
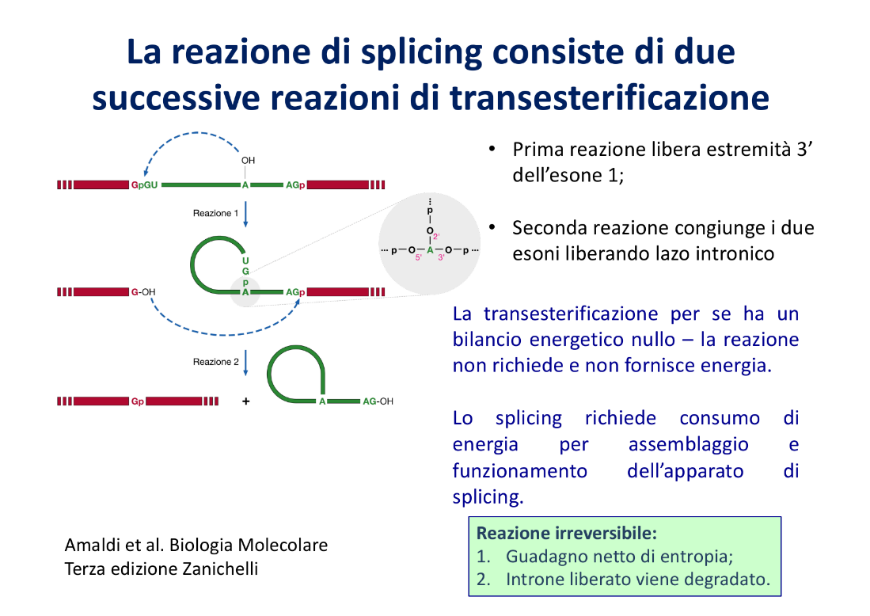
- Prima transesterificazione: Il gruppo ossidrile in posizione 2’ () dell’adenina del branch point compie un attacco nucleofilo sul fosfato in 5’ del dinucleotide GU al sito donatore. Questo rompe il legame tra l’esone a monte e l’introne, e genera un legame insolito fosfodiesterico tra la GU e l’adenina, ripiegando l’introne in una struttura a laccio (lariat).
- Seconda transesterificazione: Il gruppo ossidrile in 3’ () rimasto libero sull’esone a monte compie un attacco nucleofilo sul fosfato in 5’ del sito accettore AG, saldando covalentemente i due esoni consecutivi e liberando definitivamente l’introne sotto forma di laccio.
- Destino dell’introne: L’introne a laccio viene rapidamente degradato nel nucleo da enzimi debranching e nucleasi.
Bilancio Energetico e Termodinamica
La reazione complessiva ha un bilancio energetico neutro o favorevole:
- Le transesterificazioni consistono nel trasferimento di legami fosfodiesterici preesistenti, senza consumo netto di ATP per la catalisi chimica.
- La reazione è guidata termodinamicamente verso la direzione dei prodotti dall’aumento di entropia derivante dalla successiva degradazione dell’introne in singoli nucleotidi.
- L’accuratezza spaziale e il posizionamento dei reagenti sono coordinati dal complesso dello Spliceosoma.
🔗 Collegamenti
- Maturazione dell’RNA — 📋 fa parte di
- RNA Messaggero — 🔗 stesso meccanismo / stessa via
- Spliceosoma — 🔗 stesso meccanismo / stessa via
- Transesterificazione — 🔗 stesso meccanismo / stessa via
- Attacco Nucleofilo — 🔗 stesso meccanismo / stessa via
- Splicing Alternativo — ➡️ evoluzione / progressione
- Controllo di Qualità dell’mRNA — ➡️ evoluzione / progressione