Inizio della Traduzione
La fase di inizio è lo stadio limitante e più lento dell’intero processo di traduzione. Una regolazione così fine assicura che la sintesi proteica cominci esattamente in corrispondenza del corretto codone d’inizio, riducendo al minimo gli errori traduzionali.
In condizioni di riposo (quando non si sta traducendo), le due subunità del ribosoma si trovano in un equilibrio chimico dinamico tra la forma associata e quella dissociata:
Traduzione nei Procarioti
La subunità minore () e il tRNA iniziatore si associano al trascritto di mRNA per selezionare il primo codone AUG a monte.
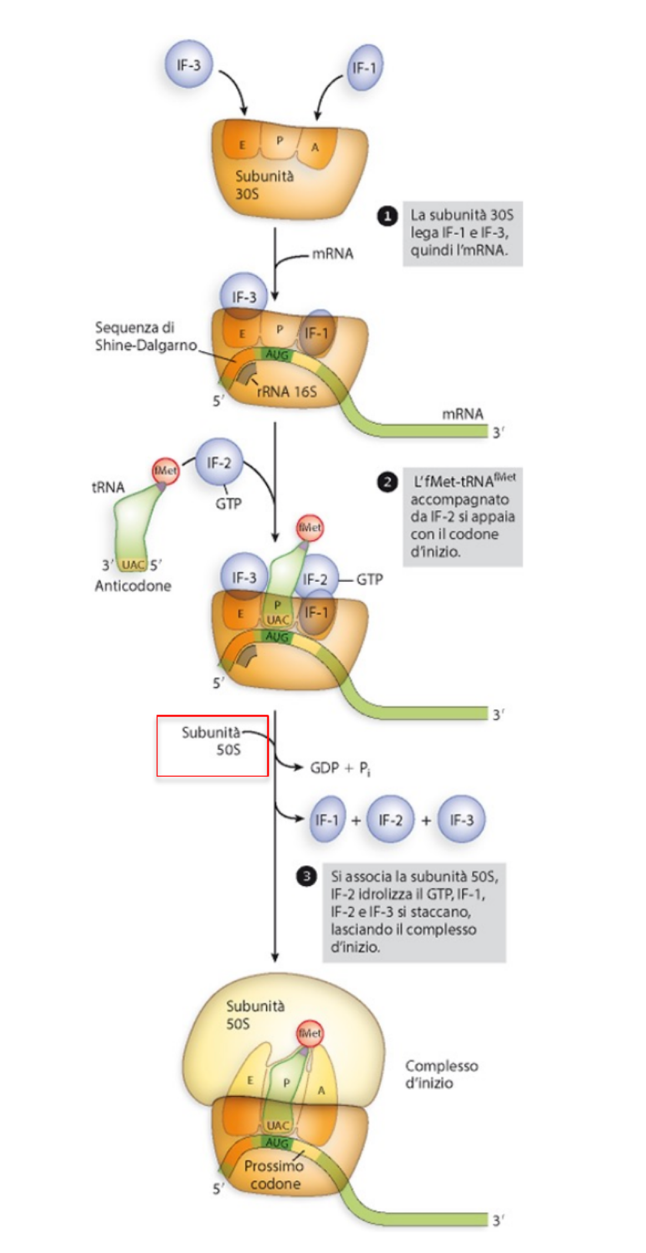
1. Mantenimento delle Subunità Dissociate e Orientamento al Sito P
Affinché possa cominciare la traduzione, le due subunità devono restare separate. Questo processo è mediato dai seguenti fattori di inizio:
- IF-3 (Initiation Factor 3): Si associa con elevata affinità all’emisito E della subunità minore . La sua funzione principale è impedire fisicamente la riassociazione prematura con la subunità maggiore .
- IF-1: Si lega all’emisito A della subunità minore, bloccando l’accesso a questa tasca.
- Di conseguenza, l’mRNA e i fattori limitano l’associazione del tRNA iniziatore unicamente al sito P.
2. Allineamento sul Trascritto: Sequenza di Shine-Dalgarno
Gli mRNA procariotici non possiedono un cappuccio al 5’. L’allineamento corretto del ribosoma avviene grazie alla sequenza di Shine-Dalgarno (SD):
- È una sequenza consenso (tipicamente
5'-AGGAGG-3') posta circa 10 nucleotidi a monte del codone d’inizio AUG. - La sequenza SD è complementare e si appaia con una regione conservata dell’rRNA 16S (sequenza
5'-CCUCCU-3') appartenente alla subunità minore del ribosoma, stabilizzando il complesso e posizionando il codone AUG esattamente nel sito P. - ⚠️ Il libro riporta invece (Capranico, p. 321): Esperimenti dimostrano che non è importante la sequenza di Shine-Dalgarno di per sé, bensì la complementarità e l’appaiamento a doppio filamento tra essa e l’rRNA 16S. (Fonte: libro)
3. Check-point Conformazionale e Idrolisi di GTP
Una volta allineato il ribosoma sul sito P:
- Giunge il tRNA iniziatore (), il quale è carico con una metionina modificata, la formilmetionina (fMet).
- Il è scortato dal fattore IF-2 legato a GTP (proteina con attività GTPasica). Il GTP qui — come per EF-Tu/EF-G nell’allungamento — funge da interruttore conformazionale: l’idrolisi è un checkpoint irreversibile che scatta solo a geometria corretta (perché GTP e non ATP: vedi EF-Tu).
- L’appaiamento corretto codone-anticodone (AUG-) e il legame stabili di IF-2-GTP agiscono da checkpoint conformazionale.
- A seguito di una corretta interazione, il GTP viene idrolizzato in GDP e fosfato inorganico (). Ciò provoca il distacco di IF-2, IF-1 e IF-3.
4. Ingresso della Subunità Maggiore
Il rilascio dei fattori d’inizio consente l’ingresso della subunità maggiore (50S), completando l’assemblaggio del ribosoma attivo () pronto per iniziare la fase di allungamento.
Traduzione negli Eucarioti
Nei sistemi eucariotici la fase di inizio risponde a un meccanismo analogo ma sensibilmente più complesso e regolato, che garantisce un’accuratezza superiore e coinvolge almeno 11 fattori di inizio eucariotici (eIF).
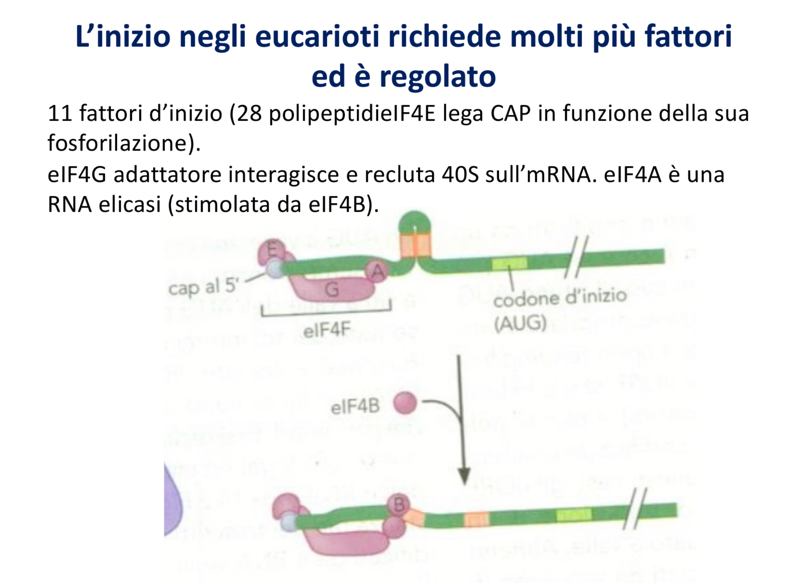
1. Riconoscimento del Cap e Risoluzione delle Strutture Secondarie
L’mRNA eucariotico non possiede una sequenza Shine-Dalgarno, ma presenta un cappuccio al 5’ (Cap):
- Il Cap viene specificamente riconosciuto dal fattore d’inizio eIF-4F (un complesso formato da tre subunità). In assenza di Cap (come in tRNA, rRNA o trascritti danneggiati), la traduzione non può iniziare.
- Poiché l’AUG d’inizio si trova più a valle, il ribosoma deve scorrere sul filamento. Per sciogliere eventuali ripiegamenti e strutture secondarie a forcina dell’mRNA incontrati durante il cammino, una delle subunità di eIF-4F possiede attività elicasica.
2. Scansione e Sequenza di Kozak
La subunità minore (che porta già associato il carico di metionina ed eIF2-GTP) effettua una scansione in direzione 5’ 3’ lungo l’mRNA a partire dal Cap. Il corretto codone di inizio AUG viene riconosciuto all’interno di una sequenza consenso specifica detta sequenza di Kozak.
3. Checkpoint e Chiusura del Ribosoma
Analogamente ai procarioti, i fattori d’inizio occupano i siti A ed E per forzare l’ingresso del metionil-tRNAi nel sito P. All’appaiamento corretto nel sito P, eIF2 idrolizza il GTP a GDP e fosfato, inducendo il distacco di tutti i fattori di inizio e l’aggregazione della subunità maggiore per formare il ribosoma completo .
⚠️ Indicazioni del docente
- Non è necessario conoscere i fattori traduzionali specifici negli eucarioti (tranne la distinzione concettuale basata sul numero di fattori complessivo e sul meccanismo di riconoscimento del Cap).
🔗 Collegamenti
- Sintesi Proteica — 📋 fa parte di
- Ribosoma — 🔗 stesso meccanismo / stessa via
- RNA Messaggero — 🔗 stesso meccanismo / stessa via
- tRNA — 🔗 stesso meccanismo / stessa via