Nucleosoma
Il nucleosoma rappresenta l’unità strutturale fondamentale e ripetitiva della cromatina negli eucarioti. Al microscopio elettronico, l’organizzazione della cromatina in fibra da 10 nm appare come una “collana di perle”, in cui i nucleosomi costituiscono i corpi sferoidali ripetuti.

Struttura
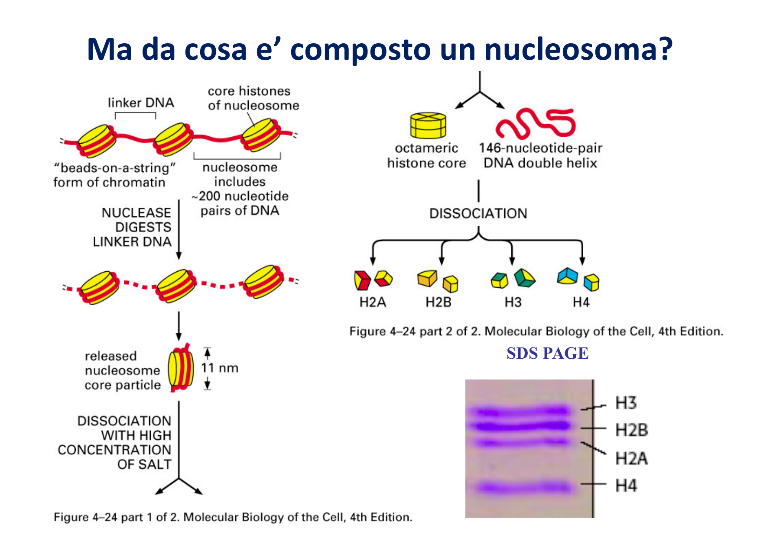
Ogni nucleosoma è composto da:
- Core Istonico: Un ottamero proteico formato da due copie per ciascuno dei quattro istoni core: H2A, H2B, H3 e H4.
- DNA Core (o nucleosomico): Un filamento di DNA a doppia elica lungo 146 coppie di basi (bp) che si avvolge esternamente attorno all’ottamero per circa 1,75 giri (o 1 e 3/4).
- DNA Linker: Il tratto di DNA nudo che connette un nucleosoma al successivo (vedi sezione dedicata sotto).
DNA Linker
Il DNA linker è il tratto di DNA a doppia elica “nudo” (non avvolto sull’ottamero) che collega un nucleosoma al successivo, dando alla fibra da 10 nm l’aspetto a “collana di perle”.
- Dimensioni: ha il diametro tipico della doppia elica B, circa 2 nm, contro i corpi nucleosomici molto più ingombranti.
- Lunghezza: è costante all’interno di uno stesso organismo ma varia tra specie differenti (in media ~20–80 bp). È la lunghezza del linker, sommata ai 146 bp del DNA core, a definire la periodicità di ripetizione del nucleosoma.
- Natura nucleotidica — prova: trattando la cromatina con endonucleasi specifiche per il DNA, il filamento di connessione di 2 nm scompare al microscopio elettronico (mentre i corpi nucleosomici restano), dimostrando che è fatto di DNA (vedi sotto, Identificazione del DNA Linker).
- Bersaglio della DNasi blanda: essendo esposto e accessibile, il linker è il primo tratto a essere tagliato da una DNasi blanda, mentre il DNA core avvolto resta protetto — è il principio dell’esperimento sull’effetto protettivo del nucleosoma.
- Sito di legame di H1: sul punto in cui il DNA entra ed esce dal nucleosoma si lega l’istone linker H1, che copre e protegge uno dei due bracci di linker e ne impone l’angolazione necessaria alla fibra da 30 nm (vedi Istone H1).
Evidenze Biochimiche e Sperimentali
La caratterizzazione molecolare del nucleosoma è stata ottenuta attraverso una serie di esperimenti di digestione enzimatica e separazione biochimica:
1. Identificazione del DNA Linker
Per dimostrare che il filamento di connessione di 2 nm fosse costituito da DNA, sono state utilizzate endonucleasi specifiche per il DNA. La digestione enzimatica ha provocato la scomparsa del filamento di collegamento osservabile al microscopio elettronico, confermandone la natura nucleotidica.
2. Natura Proteica del Nucleosoma
Il trattamento della cromatina con enzimi proteolitici (proteasi) ha degradato selettivamente i corpi sferoidali da 10 nm (i nucleosomi), dimostrandone la composizione proteica.
3. Effetto Protettivo del Nucleosoma
Trattando la cromatina purificata da nuclei lisati con una DNAsi blanda (in grado di tagliare solo il DNA linker esposto), i nucleosomi sono rimasti strutturalmente intatti. La microscopia elettronica ha rivelato la permanenza dei nucleosomi e la totale scomparsa del DNA linker.
L’integrità del DNA avvolto all’interno dei nucleosomi trattati con DNAsi è stata confermata mediante centrifugazione su gradiente di saccarosio:
- I nucleosomi hanno mantenuto inalterata la loro velocità di sedimentazione (espressa in unità Svedberg).
- Il gradiente di saccarosio ha consentito sia la purificazione dei nucleosomi dal resto dei detriti cellulari, sia l’analisi delle loro proprietà fisiche (forma, dimensioni e peso molecolare).
- Se si utilizzano DNAsi ad alte concentrazioni e in condizioni aggressive, viene digerito anche il DNA nucleosomico, lasciando isolato l’ottamero istonico, che risulta sufficientemente stabile da esistere in soluzione in forma autonoma.
4. Dissociazione e Quantificazione del DNA
L’interazione tra le proteine istoniche (cariche positivamente) e il DNA (carico negativamente per via dei gruppi fosfato) è di natura elettrostatica. L’aggiunta di elevate concentrazioni saline (alta forza ionica) a una soluzione di nucleosomi scherma le cariche elettriche dei componenti, provocando la dissociazione completa del complesso DNA-proteina.
L’analisi elettroforetica dei frammenti di DNA isolati dopo dissociazione rivela la presenza di una singola banda netta corrispondente a 146 basi, a dimostrazione del fatto che la quantità di DNA associata a ciascun nucleosoma è costante.
5. Determinazione della Stechiometria Proteica
Le proteine istoniche dissociate sono state analizzate tramite elettroforesi su gel in condizioni denaturanti (SDS-PAGE, vedi Elettroforesi su Gel per il dettaglio della tecnica). L’analisi ha rivelato la presenza di sole 4 bande proteiche distinte e di pari intensità cromatica, corrispondenti a quattro proteine di basso peso molecolare: H2A, H2B, H3 e H4.
Il ragionamento che porta all’ottamero combina due dati indipendenti:
- L’SDS-PAGE mostra 4 bande di pari intensità → gli istoni sono presenti tutti nello stesso rapporto (1 : 1 : 1 : 1). Questo dà la proporzione, ma non il numero assoluto di copie.
- Il peso molecolare reale del nucleosoma, misurato in gradiente di saccarosio, era circa il doppio del peso teorico calcolato sommando una singola copia di ciascuna delle 4 proteine (H2A + H2B + H3 + H4).
Poiché il valore reale è ~2× quello atteso per “una copia ciascuno”, e le 4 proteine stanno in rapporto 1:1:1:1, l’unico modo di far quadrare i conti è ipotizzare due copie per ogni tipo di istone: 2 × 4 = 8 proteine, cioè l’ottamero istonico. (Il peso molecolare fornisce il numero assoluto che l’SDS-PAGE da sola non poteva dare.)
Nativo vs denaturante — perché servono entrambe le tecniche: le due analisi sono complementari proprio perché trattano il campione in modo opposto. La Centrifugazione su Gradiente di Saccarosio è nativa: il complesso migra integro, quindi ne misuri il peso molecolare reale. L’SDS-PAGE è denaturante: l’SDS scinde l’ottamero nelle singole catene, perciò ne vedi i monomeri separati (4 bande) e ricavi solo il rapporto tra i tipi di istone. Integro ti dà il numero assoluto, scisso ti dà la proporzione: incrociandoli ottieni la stechiometria 2×4.
Posizione del DNA ed Avvolgimento
Per determinare se il DNA fosse avvolto esternamente o racchiuso all’interno del nucleo proteico, sono stati condotti saggi di digestione controllata:
- Se il DNA fosse stato interno, le proteasi avrebbero distrutto l’involucro proteico esterno liberando il DNA integro.
- Poiché il trattamento con DNAsi frammenta rapidamente il DNA lasciando l’ottamero proteico parzialmente protetto, è stato dimostrato che il DNA si dispone sulla superficie esterna dell’ottamero.
- Il rapporto tra la lunghezza lineare di 146 bp di DNA e la circonferenza dell’ottamero proteico determina un avvolgimento pari a circa 1 e 3/4 di giro intorno all’istone.
Meccanismo di Assemblaggio
L’ottamero istonico non si assembla in un unico passaggio, ma segue una via sequenziale regolata dalle affinità reciproche tra le subunità:
- Formazione del Tetramero H3-H4: Le proteine H3 e H4 si associano inizialmente in dimeri H3-H4, i quali polimerizzano per formare un tetramero stabile .
- Interazione Iniziale con il DNA: Il tetramero si lega per primo al DNA, formando un nucleosoma incompleto. Questa struttura iniziale è relativamente instabile e presenta un avvolgimento lasso del filamento di DNA.
- Stabilizzazione con H2A-H2B: Successivamente, due dimeri pre-formati di H2A-H2B si inseriscono ai lati del tetramero associato al DNA. L’ingresso dei dimeri H2A-H2B apporta un elevato numero di interazioni chimiche aggiuntive, stabilizzando la struttura e completando l’ottamero istonico del nucleosoma.
🔗 Collegamenti
- Istoni — 📋 fa parte di
- Proteasi — 🔬 sonda sperimentale (natura proteica, posizione del DNA)
- Compattazione della Cromatina — 📋 fa parte di
- Elettroforesi su Gel — 🔬 tecnica usata
- Centrifugazione su Gradiente di Saccarosio — 🔬 tecnica usata
- Struttura del DNA — 📋 fa parte di