Struttura del DNA
La struttura tridimensionale del DNA è stata risolta nel 1953 da James Watson e Francis Crick, i quali formularono il modello a doppia elica basandosi sui dati cristallografici a raggi X di Rosalind Franklin e sulle scoperte biochimiche di Erwin Chargaff.
Le Regole di Chargaff e la Cromatografia
Erwin Chargaff fu il primo a determinare i rapporti stechiometrici delle basi azotate nel DNA utilizzando la cromatografia su strato sottile (TLC).
Principio della Cromatografia
La cromatografia consente la separazione di biomolecole sfruttando:
- Una fase stazionaria (o immobile): Una sostanza dotata di specifiche proprietà chimiche (es. carica elettrica, idrofilia, idrofobia) depositata su un supporto rigido (come una lastra di vetro nella TLC, o resine in colonna).
- Una fase mobile (solvente): Un liquido che si muove lungo la fase stazionaria per capillarità o gravità, trascinando con sé i componenti della miscela a velocità differenti a seconda della loro affinità per le due fasi.

Esecuzione della Cromatografia su Strato Sottile (TLC)
- Chargaff isolò il DNA da vari organismi e lo sottopose a idrolisi acida per separare i singoli nucleotidi.
- Applicò la soluzione contenente i nucleotidi su una lastrina di vetro rivestita di fase stazionaria (gel di silice).
- Immerse la lastrina in una bacinella contenente il solvente (fase mobile).
- Il solvente, risalendo per capillarità, trascinò i singoli nucleotidi che migrarono a distanze diverse a seconda della base azotata contenuta, formando quattro macchie distinte.
- Utilizzando controlli positivi (nucleotidi puri con singole basi), identificò la posizione di ciascun nucleotide e calcolò la quantità di ciascuna base misurando le dimensioni e l’intensità delle macchie.


Scoperte di Chargaff
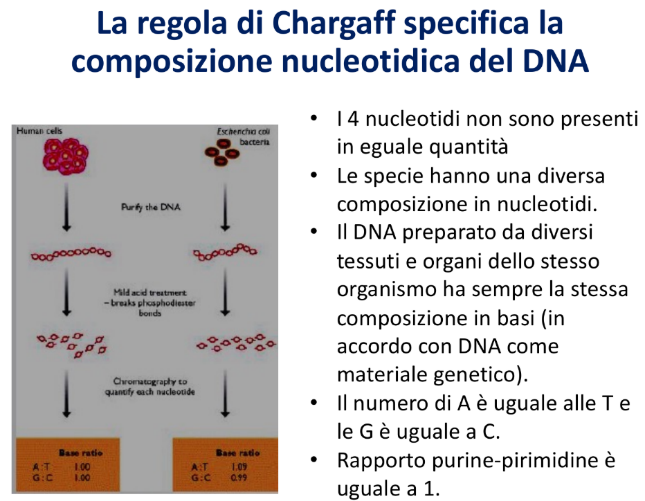
Chargaff applicò questa tecnica a DNA provenienti da diverse specie (uomo, batteri, piante, pesci), giungendo a conclusioni fondamentali:
- I quattro nucleotidi non sono presenti in quantità identiche.
- La composizione percentuale delle basi varia da specie a specie.
- La composizione del DNA umano è identica in tutti i tessuti del medesimo organismo, a testimonianza del fatto che l’informazione genetica è conservata.
- Legge di Chargaff: In qualsiasi campione di DNA a doppio filamento, la quantità di Adenina è uguale a quella di Timina () e la quantità di Guanina è uguale a quella di Citosina (). Di conseguenza, il rapporto purine/pirimidine è sempre uguale a 1 ().
Il Modello a Doppia Elica di Watson e Crick
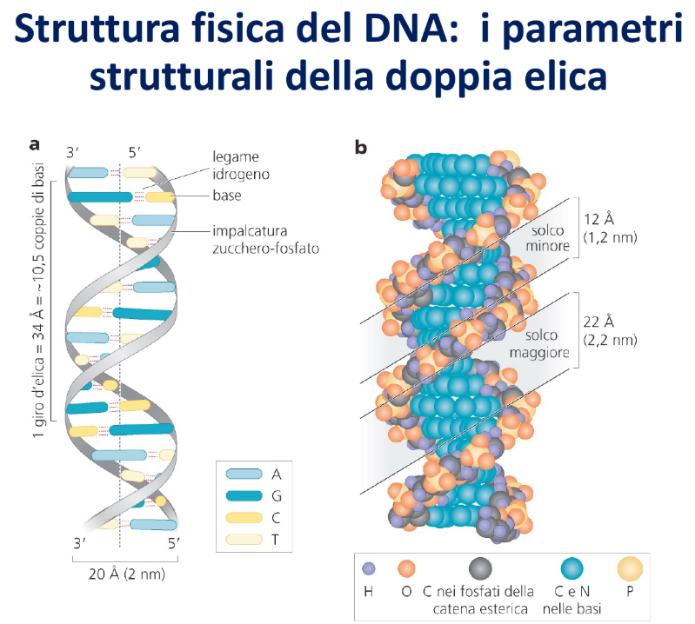
Il DNA si presenta fisiologicamente sotto forma di due filamenti polinucleotidici avvolti l’uno sull’altro. Questa conformazione a doppia elica rappresenta lo stato termodinamico più stabile della molecola (caratterizzato dalla minor energia libera possibile).
Caratteristiche Strutturali:
- Avvolgimento: I due filamenti si avvolgono a formare un’elica. (⚠️ Nota scientifica: Il DNA B è fisiologicamente una doppia elica destrogira. Tuttavia, a lezione è stato indicato un avvolgimento levogiro).
- Antiparallelismo: I due filamenti corrono in direzioni opposte. Un filamento presenta polarità (con l’estremità 5’-fosfato libera all’inizio e l’estremità 3’-OH libera alla fine), mentre il filamento complementare è capovolto e presenta polarità .
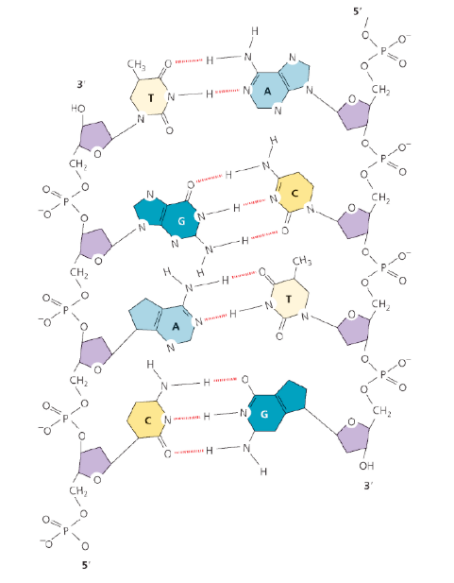
- Appaiamento Complementare: La doppia elica è tenuta unita da legami a idrogeno che si stabiliscono internamente tra le basi azotate dei filamenti opposti. L’appaiamento è altamente specifico:
- L’Adenina si appaia con la Timina mediante 2 legami a idrogeno ().
- La Guanina si appaia con la Citosina mediante 3 legami a idrogeno ().
- Diametro Costante: Poiché una purina (due anelli, ingombrante) si appaia sempre con una pirimidina (un anello, piccola), la distanza tra i due scheletri di zucchero-fosfato è costante. Il diametro dell’elica è di 2 nm (20 Å). Eventuali appaiamenti errati (mismatch) alterano localmente questo diametro creando angolature che vengono prontamente riconosciute e corrette dai sistemi di riparazione cellulari.
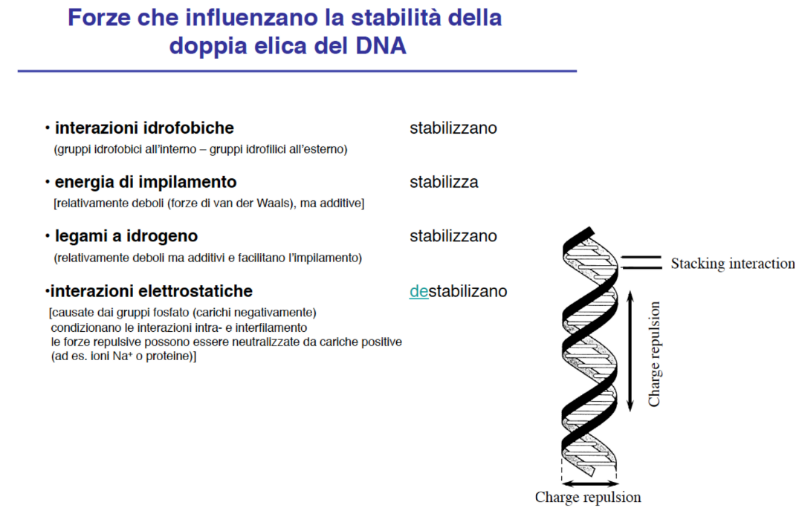
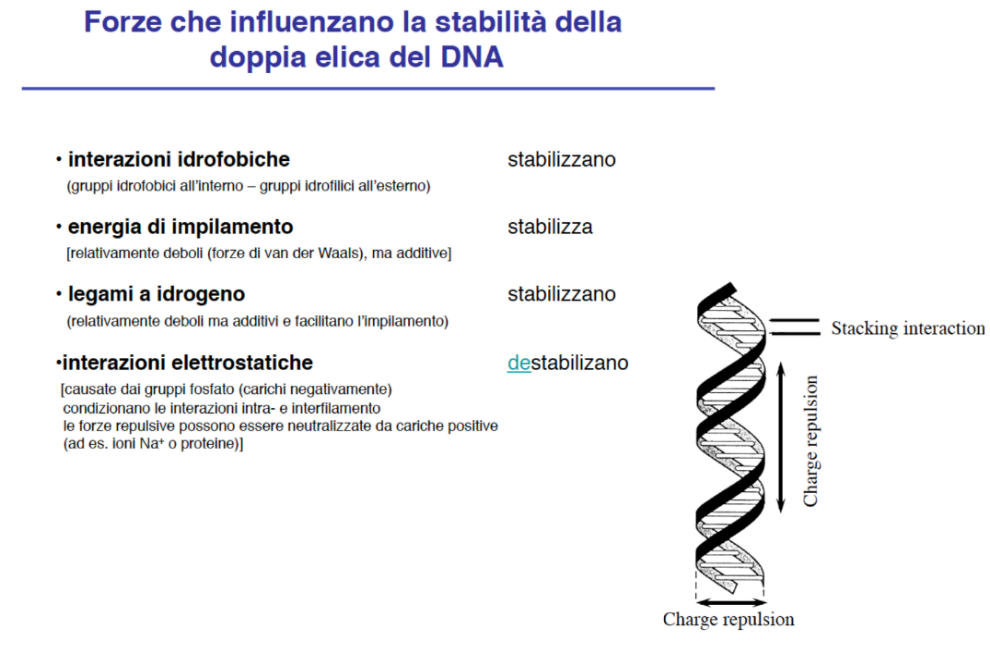
- Interazioni di Stabilità: Oltre ai legami idrogeno, la stabilità verticale della doppia elica è incrementata da interazioni idrofobiche deboli, come le forze di van der Waals e le forze di impilamento (stacking) delle basi azotate aromatiche l’una sull’altra.
- Forze Repulsive ed Effetto Schermo: Lo scheletro esterno formato dall’alternanza zucchero-fosfato porta cariche negative dovute ai gruppi fosfato. Tali cariche generano repulsione elettrostatica che tenderebbe a destabilizzare la doppia elica. Tuttavia, in ambiente cellulare acquoso, queste cariche negative sono schermate dagli ioni disciolti e dalle molecole d’acqua, impedendo la denaturazione spontanea del DNA. Le basi idrofobiche rimangono protette all’interno, mentre le porzioni idrofile e cariche (zuccheri e fosfati) sono esposte all’esterno a contatto con il solvente.
- Solchi del DNA: A causa della geometria dell’avvolgimento, sulla superficie esterna della doppia elica si formano due insenature di dimensioni differenti: il solco maggiore e il solco minore. Il solco maggiore ha una dimensione paragonabile a quella delle strutture ad -elica delle proteine; per tale motivo, le proteine regolatrici che interagiscono con sequenze specifiche di DNA (es. fattori di trascrizione) vi si legano inserendo un’elica all’interno del solco maggiore.
- Passo dell’Elica: Nella forma fisiologica prevalente (DNA B), un giro completo della doppia elica contiene circa 10.5 paia di basi.
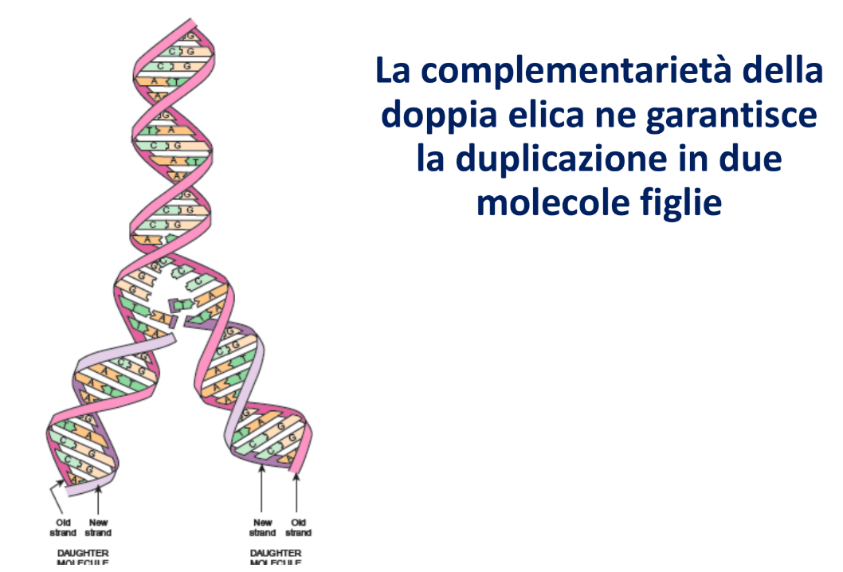
Complementarità e Replicazione
La complementarità delle basi fornisce il meccanismo molecolare per l’ereditarietà: separando i due filamenti, ciascuno di essi può fungere da stampo per la sintesi di un filamento complementare identico a quello originario, portando alla formazione di due molecole figlie identiche alla molecola madre.
Conformazioni Alternative del DNA
Oltre alla conformazione B descritta da Watson e Crick, il DNA può esistere in altre conformazioni geometriche a seconda del grado di idratazione e della sequenza nucleotidica:
- DNA A
- DNA Z (Queste conformazioni si riscontrano solo in particolari condizioni e non sono oggetto di trattazione dettagliata in questo corso).
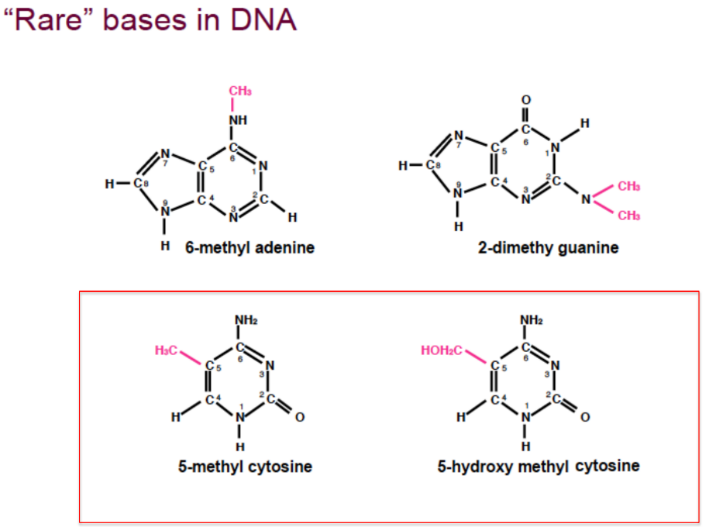
Basi Azotate Rare (Modificazioni Post-Sintetiche)
Sebbene si ritenga comunemente che il DNA contenga solo 4 basi, nei mammiferi sono presenti almeno 6 basi azotate distinte dovute a modificazioni covalenti che avvengono dopo la polimerizzazione:
- 5-metilcitosina
- 5-idrossimetilcitosina
Queste modificazioni consistono nell’aggiunta di un gruppo metilico sulla citosina ad opera di metiltransferasi. Tali basi modificate non vengono incorporate direttamente dalle DNA polimerasi (che utilizzano solo i nucleotidi canonici), ma si formano post-sinteticamente e fungono da segnali epigenetici per la regolazione e il silenziamento dell’espressione genica.
🔗 Collegamenti
- Acidi Nucleici — 📋 fa parte di
- RNA — 🔄 diagnosi differenziale / confronto
- Teoria Cellulare — 📋 fa parte di
- Rapporti Stechiometrici — 🔎 approfondimento
- DNA Levogiro e Destrogiro — 🔎 approfondimento