TCR: T-Cell Receptor
Il TCR (T cell receptor) è la struttura molecolare espressa sulla superficie dei linfociti T, finalizzata a riconoscere l’antigene e a trasdurre il segnale. Funzionalmente e concettualmente è del tutto simile al BCR: una porzione variabile che sporge dalla cellula e riconosce l’antigene (diversa su ogni clone) e una porzione costante che trasduce il segnale; la struttura di trasduzione è il CD3 (analogo di Igα/Igβ nel BCR). I linfociti T attaccano cellule infette o neoplastiche, non patogeni extracellulari.
Struttura
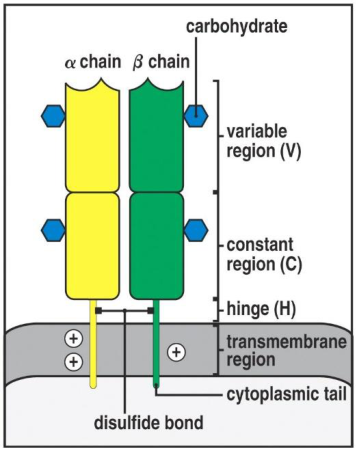
È una glicoproteina di 80.000–90.000 kDa, un eterodimero formato da due catene polipeptidiche. Differenze rispetto agli anticorpi:
- Un unico sito combinatorio (è monovalente), mentre gli anticorpi ne hanno almeno due (due Fab).
- È sempre espresso sulla superficie dei linfociti T, mai secreto (gli anticorpi possono essere secreti dalle plasmacellule). Coerente col fatto che non contrasta patogeni in circolo.
- Non subisce lo switch di classe: non cambia mai le sue regioni costanti.
L’eterodimero può essere αβ (alfa-beta) o γδ (gamma-delta); il 90–95% dei linfociti T in circolo presenta il TCR αβ. I TCR γδ sono prevalentemente tissutali e pochissimo rappresentati in circolo. Questo spiega un bias dell’immunologia: è più facile studiare il sistema immune circolante (semplice prelievo di sangue) rispetto a quello tissutale (richiede biopsie e isolamento di cellule vive).
TCR αβ: doppia restrizione
Il TCR αβ riconosce molecole MHC-HLA self (autologhe) che presentano peptidi. Vale la restrizione al self: i TCR sono attivati solo da molecole HLA self (eccezione apparente nel rigetto dei trapianti, dove riconoscono MHC non-self; va accettato di dogma). Oltre alla restrizione al self esiste la restrizione all’MHC (Restrizione MHC):
- MHC di classe I → riconosciuto solo dai linfociti CD8 (si lega all’α3); presentato dalle cellule target nucleate.
- MHC di classe II → presenta solo ai TCR dei linfociti CD4; espresso dalle APC.
CD4 e CD8 sono molecole accessorie che si associano al TCR e ne restringono la capacità di riconoscere MHC di classe I o II (Attivazione del Linfocita T).
Specificità per il complesso binario
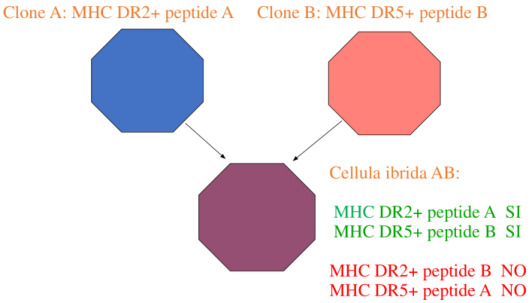
Il TCR riconosce sempre e solo il complesso binario peptide + tasca dell’MHC, mai il peptide o l’MHC isolati. Ogni tasca dell’MHC può accogliere diverse famiglie di peptidi; quindi il TCR è specifico solo per una determinata combinazione peptide X dentro la tasca di un dato MHC.
Esempio — un clone A presenta il peptide A con l’MHC DR2, un clone B presenta il peptide B con l’MHC DR5. Fondendo le due cellule, l’ibrido esprime o DR2+peptide A o DR5+peptide B: la specificità resta sempre per il peptide accolto nell’MHC. Esistono 10⁹–10¹² molecole di TCR a fronte di molte meno molecole di MHC, proprio perché la specificità è del TCR ed è diretta verso il complesso peptide+MHC.
Porzione legante l’antigene
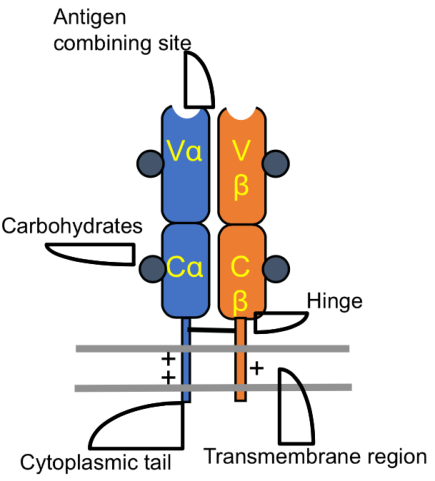
L’eterodimero αβ, come gli anticorpi, ha su entrambe le catene una porzione variabile e una costante:
- Porzione variabile (V) — distale rispetto alla cellula, lega l’MHC assieme al peptide.
- Porzione costante (C) — prossimale rispetto alla cellula.
Un ponte disolfuro tiene insieme la catena α e la catena β; una corta coda sporge nel citoplasma. La giustapposizione delle porzioni variabili α e β determina la capacità di legare i diversi peptidi accolti nell’MHC. L’analogia con gli anticorpi conferma l’ipotesi di un recettore ancestrale unico, poi differenziatosi.
🔗 Collegamenti
- CD3 — 🔗 struttura di trasduzione del segnale associata al TCR
- BCR: B-Cell Receptor — 🔄 recettore analogo sui linfociti B
- Restrizione MHC — 🔗 doppia restrizione self + classe MHC
- Geni e Diversità del TCR — ⬆️ origine della variabilità del TCR
- Attivazione del Linfocita T — ⬇️ trasduzione del segnale dopo il riconoscimento
- TCR γδ — 📋 variante tissutale dell’eterodimero
- Maturazione dei Linfociti T — ⬆️ acquisizione del TCR e selezione nel timo