Maturazione dei Linfociti T
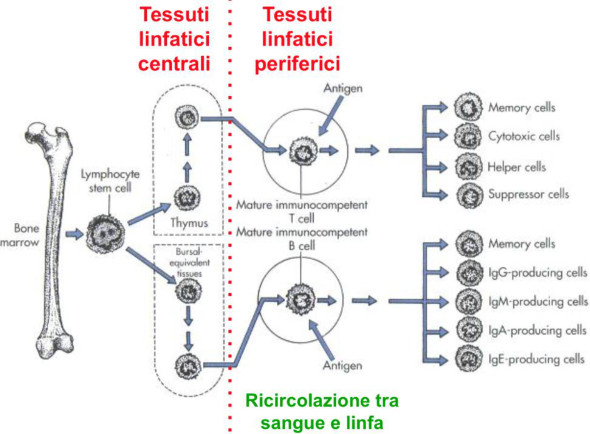
Tutte le cellule immunitarie originano nel midollo osseo dal precursore che esprime CD34. I linfociti B maturano nel midollo, mentre i linfociti T nascono nel midollo e migrano per via ematica nel timo, dove maturano. I linfociti B e T maturi lasciano gli organi linfoidi primari e non vi ritornano mai più: ricircolano tra sangue, linfa e tessuti, dove si localizzano le infezioni.
Gli organi linfoidi primari (midollo e timo) sono i siti di maturazione della linea linfoide; sono siti privilegiati, esclusi dal contatto con l’antigene e non suscettibili alle infezioni — non sono per definizione siti di risposta immune. La cellula matura li lascia e raggiunge i tessuti linfoidi secondari periferici per differenziarsi nei vari sottotipi.
Il timo
Il timo è un organo bilobato situato nel mediastino anteriore; ogni lobo è suddiviso in lobuli, ciascuno con una corticale e una midollare. Nel passaggio da corticale a midollare avvengono maturazione e selezione: i precursori arrivano dal midollo osseo come cellule pre-T e ne escono come linfociti T naive maturi, che entrano in circolo. Nel timo il 95% dei timociti muore per apoptosi.
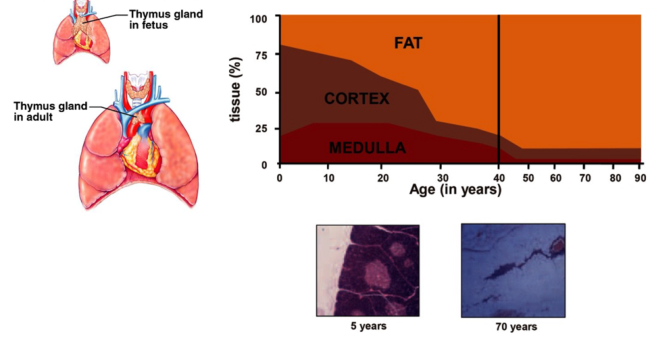
Il timo è presente fino alla pubertà, poi viene sostituito da tessuto fibroso. Non è del tutto noto dove maturino i linfociti T nell’adulto: la teoria prevalente prevede che alcuni residui timici possano espandersi all’occorrenza. Ciò si osserva nei pazienti con infezione da HIV (che hanno pochi CD4): all’inizio del trattamento, bloccando la replicazione virale, aumentano i linfociti in circolo — molti dei quali vergini — probabilmente per recupero di funzione di isole timiche persistenti nella fase post-puberale.
L’involuzione timica è mediata dalla cessazione di FOXN1, fattore di trascrizione dell’epitelio timico: dopo la pubertà smette di funzionare e l’epitelio timico degenera, sostituito da tessuto adiposo. Se FOXN1 viene riattivato nei topi anziani il timo torna a crescere e assume le sembianze di un timo giovane. Diversamente dai linfociti B (che restano in circolo 1-2 mesi e, se non incontrano l’antigene, muoiono ma vengono riformati), per i linfociti T non esiste neogenesi negli organi linfoidi primari: quelli presenti alla nascita restano per tutta la vita.
La comparsa di linfociti vergini nell’adulto è documentata in condizioni patologiche — iatrogene (es. chemioterapia prima del trapianto di midollo) o da malattia (HIV che distrugge i linfociti T). In questi pazienti, nel giro di qualche mese ricompaiono linfociti vergini nonostante il timo non sia più teoricamente funzionante. Due teorie: (1) altri organi (forse il midollo osseo) vicariano la funzione timica all’occorrenza; (2) — più accreditata — se serve, FOXN1 viene riattivato, ripristinando l’attività di isolotti timici funzionali residui che rigenerano linfociti T vergini. Questo non avviene in condizioni fisiologiche, ma solo in condizioni patologiche.
(Sezione espansa con: sbobina 9)
Tre obiettivi della maturazione
- Generazione della diversità — riarrangiamento dei geni per il TCR e generazione della variabilità del recettore.
- Acquisizione dell’autotolleranza — distruzione dei cloni che riconoscono antigeni self.
- Acquisizione della restrizione all’MHC — sopravvivono solo i linfociti il cui TCR riconosce molecole MHC self.
I linfociti T che escono dal timo sono tolleranti per il self e ristretti agli MHC self: hanno un TCR specifico per il self che presenta il non-self.
Il timo guida la restrizione MHC
La capacità di riconoscere l’MHC è guidata dal timo. Esperimenti di trapianto: un topo A timectomizzato a cui si trapianta un timo di ceppo B avrà in circolo solo linfociti ristretti a B. I linfociti che escono dal midollo potrebbero potenzialmente riconoscere qualsiasi MHC, ma la restrizione periferica è determinata dal timo e, una volta acquisita, non cambia più.
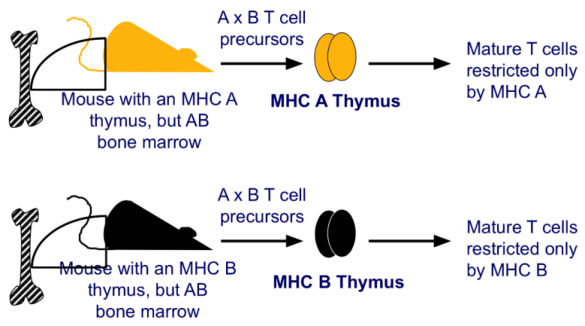
Prove sperimentali e cliniche del ruolo del timo
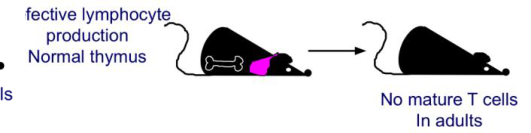
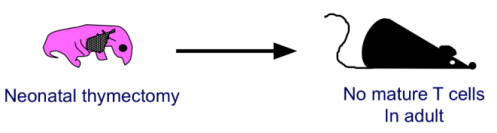
- Jacques Miller (immunologo australiano) timectomizzò topi alla nascita e scoprì che, da adulti, non producevano cellule T.
- I topi nudi (nude) sono privi di timo e di pelo: il gene che codifica per lo sviluppo timico clusterizza con i geni del pelo; anch’essi non hanno cellule T.
- La sindrome di DiGeorge è un’immunodeficienza primaria pediatrica da delezione sul braccio corto del cromosoma 22 (agenesia timica): i bambini non hanno timo né linfociti T in circolo, con prognosi grave; presentano anche difetti cerebrali di causa ignota.
Midollo osseo e timo sono entrambi indispensabili e devono collaborare: una timectomia post-natale, o un’alterazione midollare (mancano i precursori) pur con timo intatto, portano comunque all’assenza di linfociti T maturi in circolo.
Timociti: fenotipo distinto
I linfociti pre-T esprimono CD34 in quanto precursori. Entrati nel timo diventano timociti ed esprimono una famiglia di proteine diversa da quella dei linfociti T circolanti: i timociti non hanno CD3, CD4, CD8, ma marker poi soppressi durante la maturazione (CD1, CD2, CD5, CCR9, CD127, CD37). Linfociti maturi e timociti sono caratterizzati da due cluster di proteine diversi. In caso di alterazioni della funzionalità timica possono entrare in circolo timociti che hanno saltato lo sviluppo timico ed esprimono ancora questi marker.
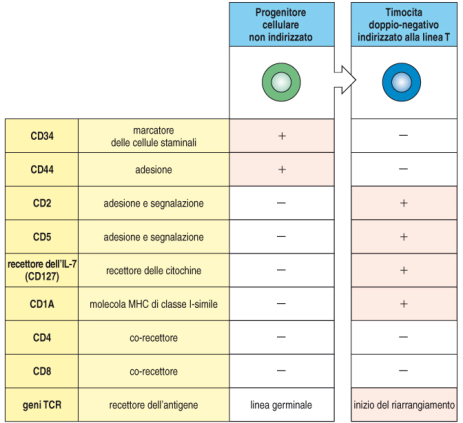
Step di maturazione
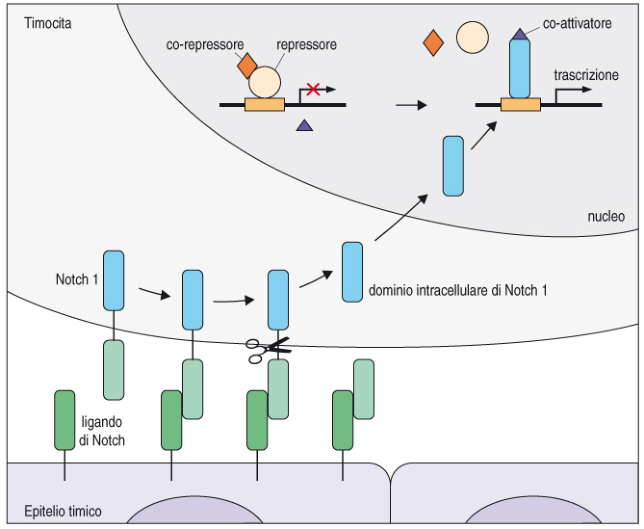
Il pre-timocita non è committed (non è ancora destinato a diventare linfocita T). Il commitment è mediato dalla proteina Notch-1, che ha un dominio extracellulare e uno intracellulare. Quando la cellula entra nel timo, il dominio extracellulare di Notch-1 si lega a uno specifico ligando sull’epitelio timico: le due subunità si scindono, quella intracellulare migra nel nucleo e attiva la trascrizione dei geni necessari alla maturazione a cellula T. Il legame Notch-1–ligando è il segnale che determina il destino della cellula.
Il primo step di maturazione è l’espressione del TCR: il timocita deve riconoscere l’MHC (il “mondo” che affronterà in periferia) e per farlo deve esprimere il TCR.
Passaggio da γδ a αβ
Nei topi (gestazione 21 giorni) e nei feti, i timociti hanno geni per tutte e quattro le catene: precocemente tutti esprimono e riarrangiano γ e δ; poi inizia il riarrangiamento di β, e alla nascita il 90–95% delle cellule possiede le catene α e β. Una proteina γ silencer silenzia la trascrizione di γ e attiva quella di β (e α); il motivo dell’attivazione intra-fetale non è noto.
La catena β neosintetizzata è trasportata nel reticolo endoplasmatico, dove si associa a una catena invariante pseudo-α. Se il legame riesce, induce la neosintesi delle catene α vere; se non riesce, la catena β viene riciclata e associata a una nuova pseudo-α, fino a 4 volte. Dopo il quarto tentativo fallito, la catena β viene distrutta.
Selezione positiva e negativa
Ciascun timocita può esprimere qualsiasi tipo di TCR. Si applicano quindi due filtri sequenziali: prima una selezione positiva (maglie ampie), poi una selezione negativa (maglie strette). Il 95% dei timociti non supera la selezione e muore.
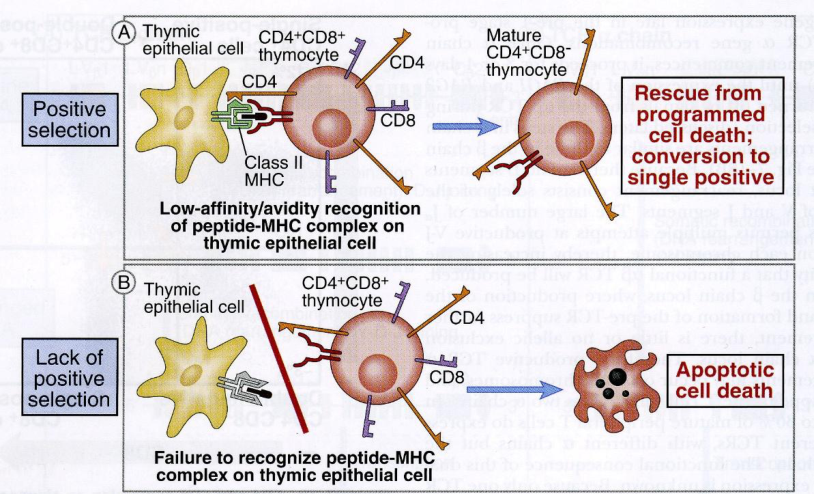
- Selezione positiva — sopravvivono i timociti il cui TCR riconosce gli MHC self espressi dall’epitelio timico → si selezionano i linfociti utili (useful).
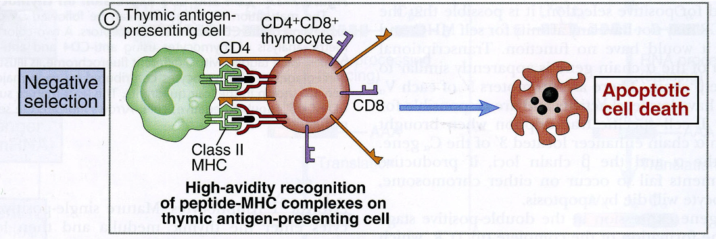
- Selezione negativa — vengono eliminati i timociti il cui TCR riconosce MHC self che presentano peptidi self → si distruggono i linfociti dannosi (harmful), che in periferia aggredirebbero i tessuti.
Nel timo sono presenti solo antigeni self (non entrano virus né batteri) e solo MHC self. Lo stesso segnale (MHC self + peptide self) salva il linfocita nella selezione positiva ma lo distrugge in quella negativa. Al termine, il repertorio è ristretto al self e tollerante verso il self: escono dal timo i linfociti con TCR specifico per il self che presenta il non-self.
Come lo stesso complesso può dare due esiti opposti
Se nel timo è presente un unico tipo di complesso (MHC self + peptide self), esso deve indurre prima la selezione positiva e poi quella negativa: i segnali devono quindi essere diversi. Sono state proposte diverse ipotesi:
- Ipotesi della densità (receptor occupancy) — è il numero di legami tra TCR e MHC a determinare l’esito: troppi legami → il timocita riconosce troppo bene il self e viene eliminato (sarebbe pericoloso in periferia); pochi legami → il timocita sopravvive.
- Ipotesi sul tempo — conta la durata del contatto tra TCR e cellula nurse: contatto troppo prolungato → il timocita riconosce troppo bene il self e viene eliminato; contatto breve → sopravvive.
- Ipotesi del segnale differenziale — il legame attiverebbe la trascrizione di un segnale biochimico qualitativamente diverso a seconda che la cellula debba sopravvivere o no. Decaduta: nessun dato la conferma.
- Ipotesi dell’affinità (la più accreditata) — se il TCR è troppo affine per l’MHC self con peptide self viene eliminato perché potenzialmente dannoso; l’affinità (forza di legame) deve essere non troppo alta. Questa ipotesi integra le prime due, dato che l’affinità deriva dalla velocità di legame (densità) e dal tempo.
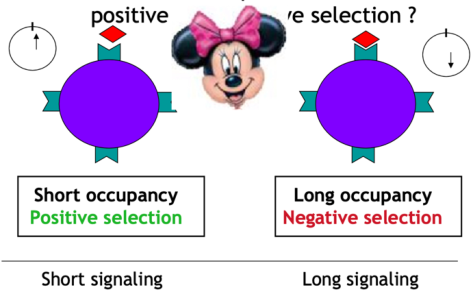
Il concetto è che si immettono in circolo linfociti T che riconoscono l’MHC, ma non troppo bene: i timociti con TCR ad alta affinità per il complesso MHC self/peptide self vanno incontro ad apoptosi (il 95% non supera il filtro timico). Un’alterazione di questa selezione è alla base di malattie autoimmuni come il lupus.
In sintesi: i timociti che entrano nel timo sono programmati a morire per apoptosi; la selezione positiva arresta lo stimolo apoptotico e induce proliferazione; se però la cellula, superato il filtro, si lega troppo bene all’MHC delle cellule nurse, lo stimolo apoptotico riparte e la cellula muore.
(Sezione espansa con: sbobina 9)
Selezione positiva: cellule nurse
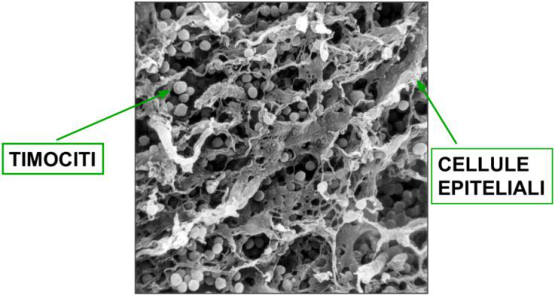
La selezione positiva permette l’acquisizione della restrizione MHC. Sull’epitelio timico si trovano le cellule nurse (cellule epiteliali) con alta densità di MHC self di classe I e II. I timociti sono programmati all’apoptosi: il segnale che la blocca è proprio il legame tra MHC self delle cellule nurse e TCR (come se ogni timocita avesse una bomba a mano innescata, disattivata solo dal riconoscimento MHC-TCR). La maggior parte dei timociti non ha un TCR capace di legare l’MHC delle cellule nurse e muore (≈90%).
Le cellule timiche producono inoltre citochine come l’IL-7, fattore solubile fondamentale per la maturazione, accanto ai fattori fisici di contatto.
🔗 Collegamenti
- TCR: T-Cell Receptor — ⬆️ il TCR è il recettore acquisito durante la maturazione
- Geni e Diversità del TCR — 🔗 riarrangiamento genico nella generazione della diversità
- Restrizione MHC — ⬇️ acquisita tramite la selezione positiva
- Maturazione dei Linfociti B — 🔄 processo parallelo nel midollo osseo
- Selezione per il Self ed Editing Recettoriale — 🔗 tolleranza centrale nei linfociti B
- Organi Linfatici — 📋 il timo come organo linfoide primario
- Marcatori CD dei Linfociti — 🔗 CD34 sul precursore, fenotipo dei timociti
- Immunodeficienza — ⬇️ sindrome di DiGeorge, agenesia timica
- Differenziazione dei Linfociti T in CD4 e CD8 — ➡️ maturazione parallela rispetto a CD4/CD8
- AIRE e Tolleranza Centrale — 🔗 rende disponibili nel timo i peptidi tissutali per la selezione negativa
- Linfociti T Regolatori (Treg) — ➡️ i Treg naturali originano dalla selezione timica