Interazioni Peptide-MHC
Promiscuità di legame
A differenza dell’anticorpo, che lega un solo antigene, ogni molecola MHC non lega un solo peptide ma intere famiglie di peptidi simili: la sua specificità è quindi meno fine (promiscuità di legame). La specificità della risposta immune non è data dall’MHC, ma dalla selettività del TCR, poiché ogni recettore riconosce un solo abbinamento peptide + MHC. Esempio: HLA-DR4 può legare sia il peptide di HBV sia quello di HCV, ma i TCR riconoscono solo il complesso HLA-DR4+HBV oppure HLA-DR4+HCV.
Ogni molecola MHC lega un solo peptide alla volta, ma può legare peptidi diversi in tempi diversi: se la tasca è piena, nessun altro peptide può entrare. In caso di infezione da più virus si crea una competizione su base di affinità.
L’MHC non è mai vuoto
Le molecole MHC sono sempre piene di peptidi: se vuote collassano. In condizioni di salute le tasche contengono proteine self derivate dal catabolismo fisiologico delle cellule. In presenza di infezione i peptidi non-self competono con quelli self, perché l’MHC non distingue tra antigene estraneo e antigene self.
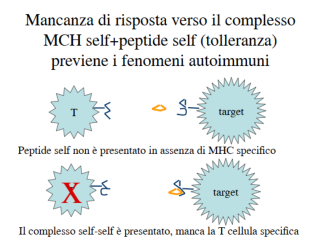
La tolleranza non deriva quindi dall’assenza di MHC, ma dall’assenza di TCR per il complesso MHC-peptide self: questo previene i fenomeni autoimmuni. Il TCR riconosce solo peptidi non-self presentati da MHC.
Cinetica del legame
L’associazione peptide-MHC è saturabile e a bassa affinità (Kd = 10⁻⁶ M), con:
- lento grado di associazione: l’antigene richiede alterazioni conformazionali (processazione) prima di potersi legare all’MHC.
- grado di dissociazione molto lento: una volta entrato nella tasca, il complesso peptide-MHC persiste a lungo sulla superficie della APC, dando modo al clone di linfociti T specifico di venire in contatto con il complesso.

Copertura allelica e lacune
La capacità di un antigene di stimolare una risposta immune dipende dalla sua capacità di essere presentato da MHC: solo gli antigeni presentati da MHC attivano i linfociti T. Alcune forme alleliche in omozigosi — HLA-B8, HLA-DR3, HLA-DQ2 — causano una risposta minima al virus dell’epatite B, perché non riescono a presentarne il peptide.
Ogni variante allelica lega solo alcuni peptidi, ma la selezione darwiniana ha fatto sì che il numero di configurazioni alleliche di ogni individuo sia sufficiente a legare quasi tutti i peptidi antigenici. La codominanza dei geni MHC e la loro immensa variabilità permettono di coprire pressoché l’intero repertorio di antigeni, pur con alcune lacune. Più sono diversi i corredi MHC di madre e padre, più vasto è il repertorio di antigeni presentabili nei figli (vantaggio evolutivo).
Uno studio su Nature condotto su 354.000 persone ha osservato una correlazione tra corredi HLA simili dei genitori e bassa statura, maggiore incidenza di patologie virali e tumorali e QI ridotto.
(Integrazione da: sbobina 7)
Il mosaico di complessi MHC-peptide
Sulla superficie di una cellula sono esposti 15-20-30 MHC diversi: poiché i geni MHC sono codominanti e ogni individuo è eterozigote, la membrana mostra un mosaico di complessi MHC-peptide diverso da cellula a cellula. Ogni molecola lega preferenzialmente una famiglia di peptidi, quindi in ogni istante è presente un campionario eterogeneo di peptidi (es. peptidi della candida, di cellule epatiche degradate, di un herpes virus cronico…).
In numeri: un’APC ha circa 100.000 molecole MHC; dividendole per le ~15-20 famiglie si ottengono circa 5.000 molecole MHC specifiche per una data famiglia di peptidi. Un linfocita T esprime 10.000 TCR, e ne bastano 15-20-50 legati per attivarlo. Ogni TCR riconosce una sola combinazione MHC-peptide, quindi solo un piccolo gruppo di MHC. Una stessa APC può attivare simultaneamente più cloni di linfociti T: in caso di infezione da più virus, si stimolano sulla stessa cellula linfociti T diversi.
🔗 Collegamenti
- MHC-HLA — 📋 funzionamento del legame peptide-MHC
- MHC di Classe I — 🔗 modalità di legame
- MHC di Classe II — 🔗 modalità di legame
- Geni dello MHC — ⬆️ la variabilità allelica determina la copertura
- Self e Non-Self — 🔗 la tolleranza dipende dall’assenza di TCR anti-self
- Co-evoluzione MHC-Patogeni — ⬇️ la pressione selettiva agisce sulla presentazione