Linfociti NK
I linfociti Natural Killer (NK) sono la seconda linea di difesa cellulare contro virus e tumori, patologie più pericolose di quelle batteriche. Sono la terza linea cellulare dell’immunità adattativa: non sono né cellule B né T, perché non esprimono né TCR né CD3 (quindi non sono T) e non hanno né CD19 né anticorpi (quindi non sono B). Esprimono il CD16.
Non maturano nel timo ma nel midollo, in sedi diverse da quelle dei Linfociti B. Sono chiamati anche LGL (Large Granular Lymphocytes) perché al microscopio ottico appaiono cellule molto grandi. Inizialmente attribuiti all’immunità innata, sono poi stati riconosciuti come parte dell’immunità adattativa per la loro complessità. Circolano nel sangue e riconoscono la cellula target in modo non specifico: riconoscono la “non salute” della cellula malata e, non avendo il TCR, non possono riconoscere antigeni presentati dalle molecole MHC. Hanno lo stesso ruolo dei citotossici: difesa contro virus e tumori.
Confronto con i citotossici
- I meccanismi d’azione sono simili a quelli dei CTL: perforina, granzimi, TNF-α, FAS-L.
- I meccanismi attivatori sono completamente diversi: gli NK non riconoscono gli antigeni presentati da MHC-I e non hanno la specificità della risposta immune.
- Hanno markers fenotipici diversi dai CD8.
Cellule LAK e tossicità da IL-2
In presenza di alte dosi di IL-2 gli NK diventano cellule LAK (Lymphokine-Activated Killer cells). Fino agli anni 2000 si è investito molto nell’immunoterapia dei tumori basata su queste cellule: si estraevano, purificavano ed espandevano in vitro gli NK di un paziente tumorale in presenza di alte dosi di IL-2, trasformandoli in cellule LAK con cui si sperava di curare il tumore. Le cellule LAK si sono però rivelate molto tossiche e molti pazienti morivano: la terapia LAK è stata abbandonata.
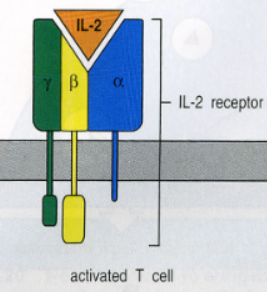
La tossicità dipende dal recettore per IL-2, che esiste in due forme:
- Dimerico (catene βγ): bassa affinità, presente sulle cellule a riposo.
- Trimerico (βγ + catena α, CD25): alta affinità, presente sulle cellule attivate; basta una bassa quantità di IL-2 per attivare i linfociti.
Gli NK non trascrivono mai il gene per la catena α, quindi presentano sempre il recettore a bassa affinità. Per attivarli e formare cellule LAK servono perciò alte dosi di IL-2, che provocano tossicità e cytokine storm (la stessa tempesta citochinica che ha ucciso molti pazienti COVID): attivazione eccessiva del sistema immune, produzione abnorme di citochine e tossicità. Questo è il motivo del fallimento della terapia LAK.
Meccanismi di attivazione
ADCC (Antibody-Dependent Cell-mediated Cytotoxicity)
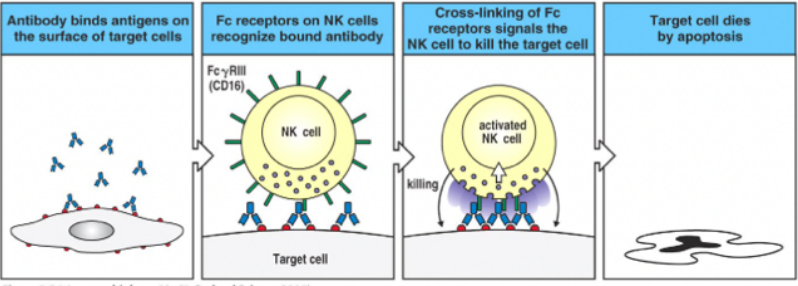
Prima ipotesi storica, per molti anni ritenuta l’unico meccanismo. Si producono anticorpi specifici per l’antigene tumorale o virale espresso sulla cellula target: l’anticorpo lega con la porzione variabile l’antigene sul target e con la porzione Fc il CD16 dell’NK. In questo modo l’anticorpo avvicina l’NK al target, l’NK si attiva e lo distrugge.
Missing self / recettori inibitori
Vent’anni dopo si è capito che gli NK hanno recettori che riconoscono MHC-I, presente su tutte le cellule sane. MHC-I lega il recettore KIR sull’NK. Gli NK controllano a random che le cellule esprimano MHC-I: se MHC-I è presente, il KIR vi si lega e sopprime l’attivazione dell’NK (KIR è un recettore soppressore).
Una cellula malata, tumorale o infettata, produce IL-10 come meccanismo di evasione dalla risposta immune: l’IL-10 riduce l’espressione di MHC-I, impedendo il legame con KIR e quindi attivando l’NK. È un meccanismo mediato dall’assenza del self (mancanza di MHC-I), non dalla presenza di un segnale di pericolo. Le cellule sane, con alta densità di MHC-I, non vengono uccise; i target sono soprattutto le cellule infette e le cellule tumorali metastatiche.
Nell’arms race tra patogeni e sistema immune, virus e tumori hanno sviluppato la capacità di sopprimere MHC-I per sfuggire ai citotossici; gli NK rappresentano il meccanismo compensatorio del sistema immune contro questa evasione.
Recettori attivatori
Gli NK hanno anche recettori attivatori: NKp30, NKp44, NKp46. Questi legano proteine come CD112 e CD155, espresse solo su cellule malate, attivando l’NK.
Il bilancio tra i due segnali determina l’attivazione:
- Cellula sana: alta densità di MHC-I (legame con KIR → soppressione) e nessuna proteina per i recettori attivatori → NK inattivo.
- Cellula stressata: esprime le proteine per i recettori attivatori ma mantiene alta densità di MHC-I → attivazione molto contenuta.
- Cellula pericolosa: forte riduzione di MHC-I e forte aumento delle proteine attivatorie → forte attivazione dell’NK.
Diverse patologie presentano varianti alleliche dei KIR e delle proteine attivatorie, con maggiore o minore attivazione degli NK.
🔗 Collegamenti
- Linfociti T Citotossici (CTL) — 🔄 stesso ruolo effettore (anti-virus, anti-tumore), meccanismi d’azione condivisi ma attivazione diversa
- MHC di Classe I — 🔗 il suo riconoscimento via KIR sopprime l’NK (missing self)
- Funzioni degli Anticorpi — 🔗 l’ADCC sfrutta la porzione Fc dell’anticorpo legata al CD16
- Linfociti NKT — 🔄 popolazione ibrida tra NK e citotossici