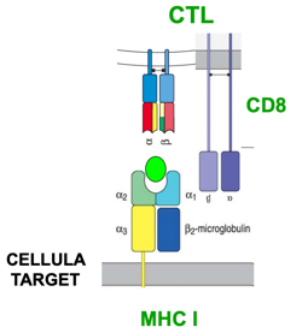
Linfociti T Citotossici (CTL)
I linfociti T citotossici (CTL) sono il meccanismo effettore dell’immunità cellulare contro i patogeni intracellulari. Esprimono CD3 e sono CD8+, riconoscono gli antigeni presentati da MHC di Classe I e, una volta maturati, inducono la lisi delle cellule infette o degenerate in senso neoplastico.
Il braccio cellulo-mediato difende dai patogeni a localizzazione intracellulare: virus, cellule degenerate in senso neoplastico, batteri con ciclo di replicazione intracellulare (es. Mycobacterium tuberculosis) e protozoi con fasi di replicazione intracellulari (es. plasmodio della malaria). Questi meccanismi sono mediati dai linfociti citotossici, dai Linfociti NK e dai macrofagi attivati. Si distingue dalla difesa verso i patogeni extracellulari (batteri extracellulari, particelle virali libere, fasi extracellulari dei protozoi), affidata invece ad anticorpi, complemento e fagociti.
I tre meccanismi effettori dell’immunità cellulare citotossica sono affidati a tre popolazioni che agiscono con modalità simili: i linfociti citotossici (CD3+ CD8+), i Linfociti NK (CD16+) e i Linfociti NKT.
Origine e frequenza
I CTL si sviluppano nel timo, come tutti i Linfociti T, a partire dai timociti double positive (CD4 e CD8), che diventano single positive CD8 in seguito al legame con MHC-I sulle cellule nurse (Differenziazione dei Linfociti T in CD4 e CD8). In condizioni di salute rappresentano il 35% dei linfociti T circolanti: un’alterazione di questo rapporto è segno patognomonico di infezione da HIV, che distrugge i CD4 e non i CD8, facendone aumentare la percentuale. Una piccola quota di citotossici non CD8 è documentata solo nel topo; nell’uomo i citotossici sono CD8.
Prima dell’incontro con l’antigene i CD8 circolano a bassa frequenza (5-6 cloni per citotossico vergine) sotto forma di linfociti pre-citotossici, privi di capacità litica. Dopo il riconoscimento dell’antigene i cloni si espandono diventando milioni e acquisiscono capacità litica e apoptotica.
Maturazione e attivazione
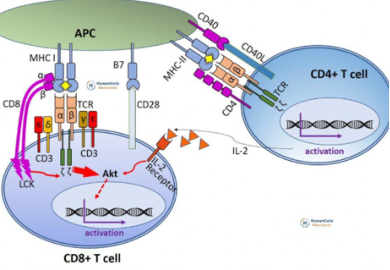
La maturazione del linfocita pre-citotossico a cellula litica è secondaria alla produzione di citochine da parte dei Th1. Il modello classico prevede un cluster di tre cellule:
- La cellula infetta, che presenta antigeni in MHC-I e anche in MHC-II (alcuni antigeni sfuggono alla cellula e vengono ricatturati e reinseriti nel pathway di classe II). Presenta prima gli antigeni di classe I al linfocita CD8.
- Il linfocita CD8 pre-citotossico, attivato dalle citochine prodotte dal CD4.
- Il linfocita Th1 (CD4 differenziato in senso Th1, che riconosce gli antigeni presentati da MHC-II): produce le citochine di tipo 1 (IL-2, IFN-γ, IL-12). Queste attivano i citotossici, armano i macrofagi aumentandone la capacità microbicida e inducono la differenziazione in senso IgG3 (anticorpi opsonizzanti che facilitano i fagociti); nel complesso stimolano l’infiammazione e l’attivazione del sistema immune.
Difficoltà del modello classico e teorie alternative
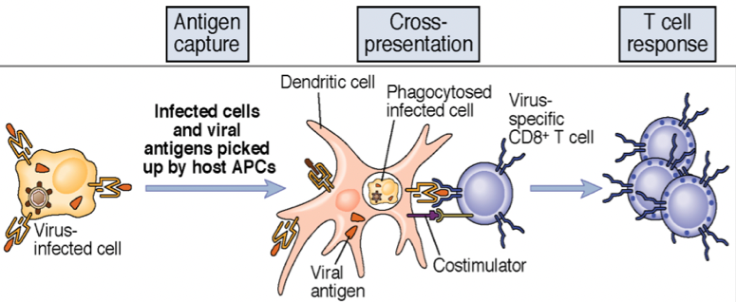
Il cluster di tre cellule presenta una difficoltà teorica: sia il CD4 sia il CD8 devono riconoscere lo stesso patogeno, ma alcuni peptidi virali sono presentati in classe I e altri in classe II. Nell’esempio dell’epatite, il cluster comprende la cellula infetta dal virus, il CD4 Th1 (che riconosce i peptidi in classe II) e il CD8 (che riconosce i peptidi in classe I), attivato dalle citochine del CD4.
Sono stati proposti meccanismi che spiegano la doppia presentazione:
- I virus hanno una fase extracellulare prima di entrare nella cellula: in questa fase sono endocitati e presentati in classe II; quando infettano la cellula entrano nel pathway di classe I. Si forma il cluster di tre cellule.
- La cellula infettata dal virus viene fagocitata da una cellula dendritica: gli antigeni vengono presentati in classe I, mentre la fase extracellulare del virus consente anche la presentazione in classe II. Anche qui cluster di tre cellule.
- La spiegazione più semplice e probabilmente reale: le dendritiche infettate presentano solo in classe I ma secernono citochine (soprattutto IL-2) che inducono maturazione, attivazione e differenziazione del CD8. In questo caso il cluster di tre cellule non serve: si prescinde dal CD4.
Caratteristiche del CTL maturo
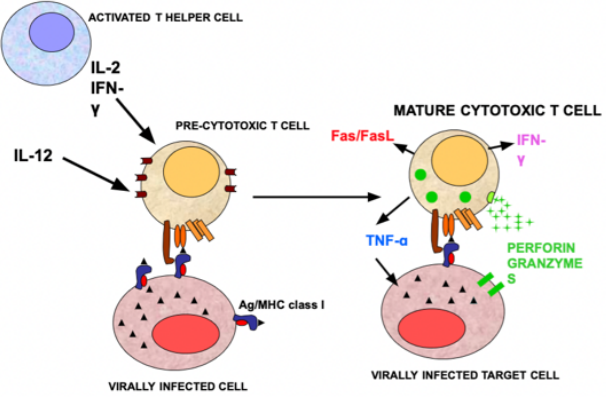
In seguito alla presentazione dell’antigene in classe I (pre-stimolo) e alla produzione di citochine (IL-12, IFN-γ) da parte del Th1 o della dendritica, il pre-citotossico matura in CTL litico, che acquisisce:
- Produzione di perforina, proteina che forma buchi sulla cellula target.
- Produzione di granzimi, che inducono l’apoptosi della cellula target.
- Espressione di FAS-L (ligando) che, legandosi a FAS sulla cellula target, ne induce l’apoptosi.
- Capacità di produrre TNF-α, che induce l’apoptosi della cellula target.
- Capacità di produrre IFN-γ, che aumenta l’espressione di MHC-I incrementando l’efficienza del sistema.
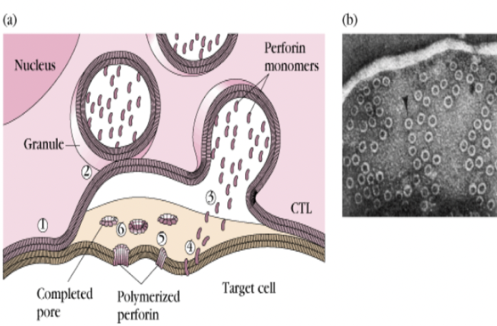
Perforina e granzimi sono contenuti in granuli citoplasmatici; il TNF-α è secreto.
Meccanismi di distruzione del target
Il CTL maturo uccide la cellula per lisi osmotica o per apoptosi. I meccanismi sono altamente selettivi (colpiscono solo la cellula target che espone l’antigene di classe I virale o neoplastico) e sinergici, non alternativi:
- Meccanismo litico: mediato dalla perforina.
- Meccanismo apoptotico:
- Solubile: mediato dal TNF-α.
- Recettoriale: il FAS-L sul citotossico si lega a FAS sulla cellula target inducendone l’apoptosi.
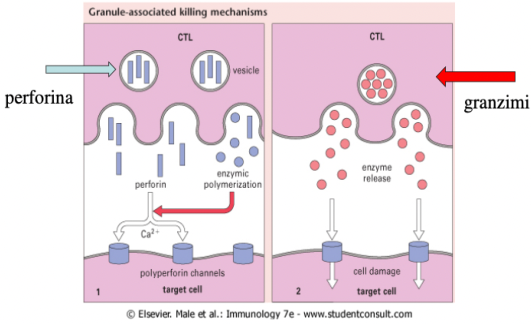
- Iniettabile: le perforine formano buchi sulla superficie del target, attraverso i quali entrano i granzimi che attivano le caspasi, inducendo la frammentazione del DNA.
Letal hit
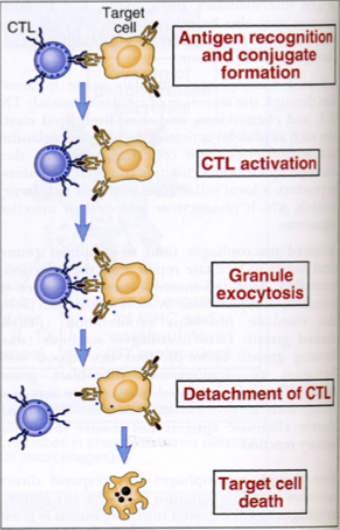
Il contatto tra le due cellule attiva il linfocita citotossico; segue il letal hit (colpo letale), il fenomeno cellulare che induce la distruzione del target. Dopo il letal hit il CTL si stacca e la cellula target muore; il CTL può uccidere molte cellule in sequenza, perché il virus infetta molte cellule contemporaneamente.
Nella zona di contatto i granuli intracitoplasmatici (perforina e granzimi) migrano all’interfaccia tra le due cellule. La perforina viene rilasciata sotto forma di monomeri che, in presenza di alte concentrazioni di calcio extracellulare, si infilano nella membrana del target e polimerizzano formando i buchi. La perforina forma buchi solo sulla cellula target perché il CTL possiede un enzima che ne impedisce la polimerizzazione sulla propria membrana, evitando l’autodistruzione. Attraverso i buchi entrano i granzimi che attivano le caspasi e la frammentazione del DNA. In aggiunta, il FAS-L del CTL si lega a FAS (presente su tutte le cellule), attivando ulteriormente le caspasi.
Questa ridondanza di meccanismi litici e apoptotici garantisce l’eliminazione del pericolo: la cellula neoplastica viene uccisa prima di formare un tumore clinico, la cellula infetta prima che l’infezione diventi letale per l’ospite.

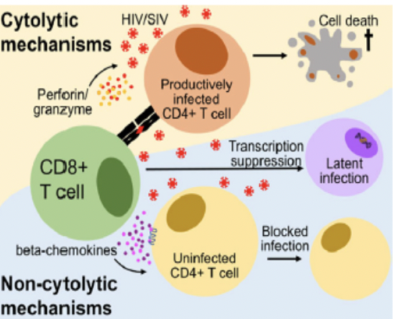
Controllo della replicazione virale tramite fattori solubili
I meccanismi con cui i CD8 ostacolano l’infezione, oltre alla lisi diretta, sono stati scoperti studiando l’HIV (epidemia dal 1983, terapia efficace dal 1996). Nel caso dell’HIV il CD8 uccide con meccanismi litici e apoptotici il linfocita CD4 infettato: questo contribuisce alla progressione dell’AIDS, perché il CD8 svolge il suo lavoro ma elimina i CD4.
Oltre alla lisi, il CD8 può:
- Sopprimere la replicazione del virus nelle cellule infette grazie alla produzione di fattori solubili non ancora identificati.
- Proteggere le cellule non ancora infette grazie alla produzione di chemochine β.
- Produrre alte quantità di IFN-γ, che amplifica l’espressione di MHC-I aumentando l’efficienza del sistema.
🔗 Collegamenti
- Sottotipi Funzionali dei Linfociti T CD4+ — ⬆️ i Th1 forniscono le citochine che attivano i CTL
- MHC di Classe I — 🔗 presenta gli antigeni riconosciuti dai CD8
- Linfociti NK — 🔄 stesso ruolo effettore (anti-virus e anti-tumore), attivazione diversa
- Linfociti NKT — 🔄 popolazione citotossica tissutale
- Differenziazione dei Linfociti T in CD4 e CD8 — ⬆️ origine timica dei CD8