Regolazione del Complemento
I circuiti regolatori spengono il complemento attivato, inibendolo. È necessario perché molte proteine del complemento sono estremamente reattive: vanno spente sia al termine della loro funzione (patogeno eliminato), sia quando il complemento si attiva per uno scorretto riconoscimento di un antigene self.
Quasi tutte le proteine regolatorie sono codificate da geni omologhi del cromosoma 1, sono omologhe tra loro e hanno funzione inibitoria (unica eccezione la properdina, che invece promuove la cascata). La loro mancanza causa diverse patologie.
C1 INH — inibizione della via classica
Inibitore di tutte le serin-proteasi, blocca la via classica agendo su C1. Quando una singola IgG (non legata al patogeno) richiama il tetramero C1r-C1s, il substrato preferenziale del tetramero diventa C1 INH anziché C4: C1 INH si lega al tetramero e lo stacca da C1q, spegnendo il segnale e arrestando la cascata. C1 INH blocca anche la cascata della coagulazione (vedi Angioedema Ereditario).
Blocco della C3 convertasi
Alcune proteine si legano a C3b o C4b impedendo l’assemblaggio degli altri componenti della C3 convertasi:
- CR1 (proteina di membrana);
- MCP (proteina di membrana);
- DAF (proteina di membrana);
- C4bp (proteina plasmatica, specifica per via classica e lectinica);
- fattore H (proteina plasmatica, specifica per via alternativa).
Le membrane dei mammiferi sono ricoperte da acido sialico: quando una proteina del complemento riconosce per errore un antigene self, l’interazione con l’acido sialico ne aumenta l’affinità per le proteine regolatorie piuttosto che per quelle del complemento, così vengono richiamate le regolatorie.
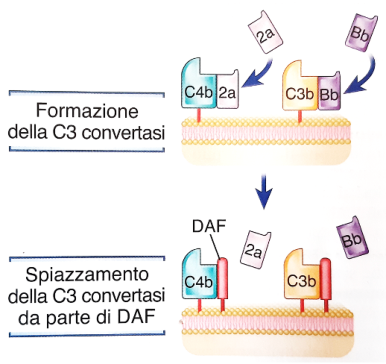
Esempio (DAF): C3b/C4b riconoscono l’antigene self sul tessuto dell’ospite ricco di acido sialico; arriva DAF e si forma un legame covalente DAF-C4b che impedisce l’avanzamento della cascata. Lo stesso avviene tra fattore H e C3b.
Degradazione proteolitica di C3b e C4b
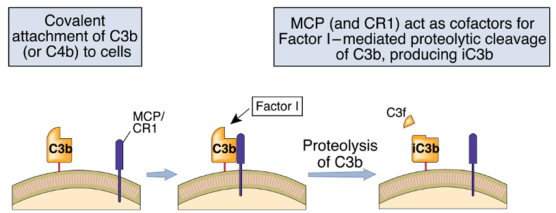
Quando C3b/C4b si legano per errore ad antigeni self, si legano a una proteina regolatoria (es. MCP) che richiama il fattore I. Il fattore I taglia ulteriormente C3b producendo i frammenti che vengono riconosciuti dai recettori del complemento (C3b inattivo, C3dg, C3d) e che permettono l’opsonizzazione.
Inibizione della formazione del MAC
Due proteine inibiscono la formazione del MAC a livello della via effettrice:
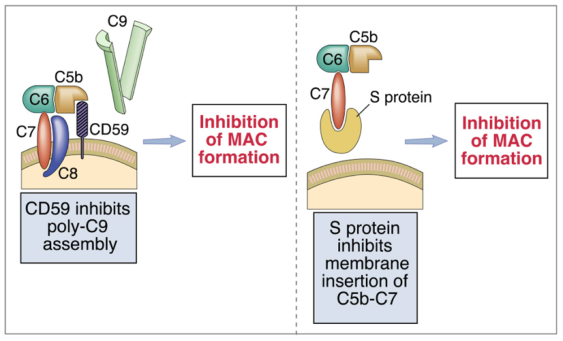
- CD59: proteina di membrana, agisce a valle. Quando il complesso C5b-C6-C7-C8 si inserisce per errore nel tessuto dell’ospite, CD59 lo riconosce, vi si lega covalentemente e impedisce il legame con C9, bloccando la formazione del MAC.
- proteina S: proteina plasmatica, agisce a monte. Quando C7 (nel complesso C5b-C6-C7) sta per inserirsi nel tessuto dell’ospite, la proteina S si lega a C7 e ne impedisce l’inserimento.
Ancoraggio delle proteine regolatorie di membrana
Le proteine regolatorie di membrana (CD59 e DAF) sono piccole e hanno un segmento idrofobo troppo corto per legarsi direttamente alla membrana: necessitano di un residuo di glicofosfatidilinositolo (GPI) presente in membrana, a cui si legano indirettamente. In alcune patologie manca l’enzima che media questo legame, perciò le proteine non sono presenti in membrana (vedi Emoglobinuria Parossistica Notturna (EPN)).
🔗 Collegamenti
- Sistema del Complemento — 📋 spegnimento della cascata
- Via Classica del Complemento — ⬇️ C1 INH blocca C1
- Via Alternativa del Complemento — ⬇️ fattore H blocca la C3 convertasi
- Via Effettrice e Complesso MAC — ⬇️ CD59 e proteina S inibiscono il MAC
- Angioedema Ereditario — ⬇️ deficit di C1 INH
- Emoglobinuria Parossistica Notturna (EPN) — ⬇️ assenza di CD59/DAF per deficit di GPI
- Deficit delle Proteine del Complemento — ⬇️ deficit di fattore I e fattore H