Via Classica del Complemento
Via di attivazione del complemento che richiede la presenza di anticorpi che medino l’interazione tra l’antigene del patogeno e le proteine del complemento. L’attivazione dipende solo dagli anticorpi: appartiene quindi all’immunità umorale.
Componenti
Le proteine specifiche della via classica sono C1, C4 e C2, presenti come zimogeni nel torrente circolatorio. A queste si aggiunge C3, condivisa con le altre due vie.
C1 è un complesso pentamerico costituito da:
- una subunità C1q, che riconosce gli anticorpi e dà inizio alla cascata;
- un tetramero formato da due copie del dimero C1r-C1s, serin-esterasi con attività proteolitica che attivano le proteine successive della cascata.
C1q ha forma a ombrello, con bracci radiali che legano C1s e C1r e teste globulari che si legano alle porzioni Fc degli anticorpi.
Attivazione di C1 e regolazione interna
C1q per attivarsi necessita di anticorpi legati ai patogeni. Se C1q incontrasse un singolo anticorpo libero (non legato a patogeni) avvierebbe la cascata con il rischio di danneggiare un tessuto self vicino (tessuto vascolare). Per evitarlo, esiste un sistema di regolazione interna: C1 si attiva solo se C1q riconosce almeno due porzioni Fc. Due Fc vicine indicano con maggiore probabilità anticorpi legati allo stesso patogeno.
Le classi anticorpali che attivano la via classica, dopo il riconoscimento di un antigene non-self multivalente, sono IgM e IgG:
- servono almeno 2 IgG per attivare una C1;
- una singola IgM pentamerica possiede 5 porzioni Fc, quindi ne basta una. Quando l’IgM non è legata all’antigene le Fc sono nascoste e C1 non può legarsi; legandosi all’antigene l’IgM cambia conformazione ed espone le 5 Fc. Una IgM può così attivare 2 molecole di C1.
Cascata enzimatica
Quando C1q lega le Fc cambia conformazione ed espone siti che attirano C1r e C1s, che si assemblano nel tetramero. C1r si attiva per prima e cliva C1s, attivandola.
C1s attivata cliva C4 in due frammenti:
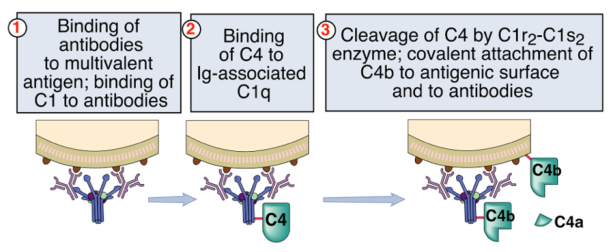
- C4a: frammento piccolo, privo di attività enzimatica; torna in circolo con funzione proinfiammatoria, chemiotattica e di flogosi.
- C4b: frammento grande con attività enzimatica. Alcune molecole di C4b restano legate al complesso in formazione, altre funzionano da opsonine (ricoprono il patogeno e richiamano i fagociti).
La presenza di C4b nel complesso richiama C2, anch’essa clivata da C1s:
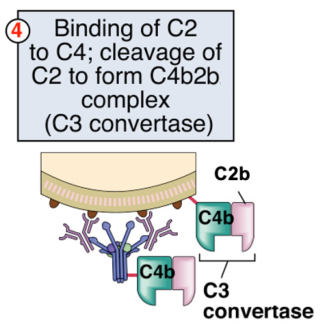
- C2a: frammento piccolo con funzione di flogosi e chemiotassi; torna in circolo.
- C2b: frammento grande con attività enzimatica, resta legato a C4b.
Il complesso C4b + C2b è la C3 convertasi della via classica, che cliva C3:
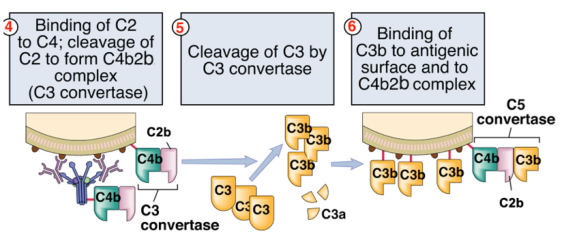
- C3a: frammento piccolo, torna in circolo con azione flogistica.
- C3b: funziona da opsonina, oppure resta legato al complesso.
Il C3b che resta legato forma con C4b e C2b la C5 convertasi (C4b + C2b + C3b), che cliva C5 (ultima proteina della via classica):
- C5a: rilasciata in circolo; è un’anafilotossina molto potente (induce rilascio di istamina, forte azione flogistica), più potente di C4a e C3a.
- C5b: frammento con funzione enzimatica, su cui convergono tutte e tre le vie. Dà inizio alla Via Effettrice e Complesso MAC.
Cross-talk tra le vie
Il C3b della via classica è estremamente reattivo: una volta riconosciuto l’antigene può attivare la Via Alternativa del Complemento; viceversa il C3b prodotto dalle vie alternativa e lectinica può attivare la cascata della via classica.
🔗 Collegamenti
- Sistema del Complemento — 📋 via di attivazione
- Classi di Immunoglobuline — ⬆️ IgM e IgG attivano C1q
- Funzioni degli Anticorpi — 🔗 attivazione del complemento via Fc
- Via Lectinica del Complemento — 🔄 cascata analoga (MASP ≈ C1r/C1s)
- Via Effettrice e Complesso MAC — ⬇️ convergenza su C5b
- Regolazione del Complemento — 🔄 C1 INH e blocco della C3 convertasi