Cicline e Chinasi Ciclina-Dipendenti
Le cicline e le chinasi ciclina-dipendenti (CDK) costituiscono il motore molecolare che regola la progressione e la transizione ordinata della cellula attraverso le fasi del ciclo cellulare.
Interazione e Concentrazione
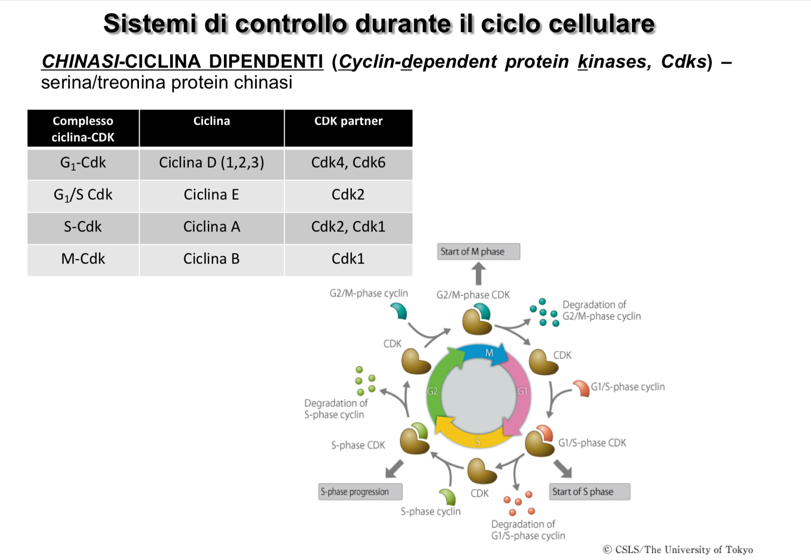
- Chinasi Ciclina-Dipendenti (CDK): Enzimi responsabili della fosforilazione di proteine bersaglio chiave per avviare le diverse fasi del ciclo. La loro concentrazione intracellulare è costante durante l’intero ciclo cellulare, ma rimangono inattive dal punto di vista catalitico se non sono associate a una ciclina.
- CDK dei mammiferi: Le principali sono CDK1, CDK2, CDK4 e CDK6.
- Specificità: Gli accoppiamenti non sono esclusivi; ad esempio, CDK1 può legarsi sia alla Ciclina B che alla Ciclina A.
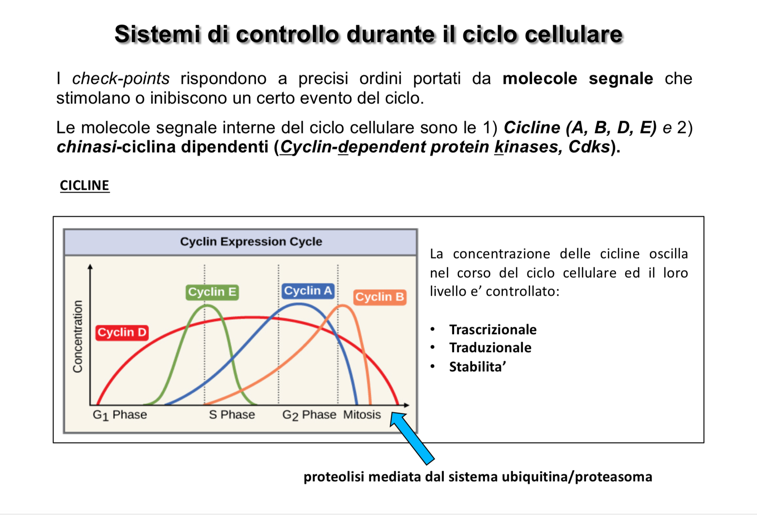
- Cicline: Proteine regolatrici prive di attività enzimatica intrinseca, ma indispensabili per legare e attivare le CDK. La loro concentrazione varia e oscilla periodicamente a seconda della fase del ciclo cellulare.
Biosintesi e Degradazione delle Cicline
La regolazione dei livelli di cicline è estremamente dinamica e governata da due processi opposti:
- Sintesi (Accumulo): Viene promossa al momento del bisogno attraverso l’attivazione della trascrizione e della traduzione dei rispettivi geni.
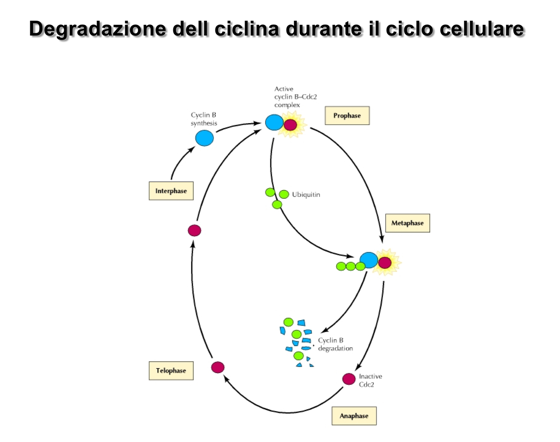
- Degradazione (Crollo): Al termine della loro finestra di attività, le cicline (in quanto proteine solubili citoplasmatiche) non vengono degradate nei lisosomi, ma attraverso il sistema del proteasoma.
- Meccanismo: La disattivazione del complesso inizia con la fosforilazione della ciclina, che funge da segnale per la sua poliubiquitinazione. I corpi poliubiquitinati vengono riconosciuti dal proteasoma, che ne promuove la rapida degradazione proteolitica, liberando la CDK per future associazioni. Per il meccanismo generale di marcatura e degradazione vedi ubiquitinazione e proteasoma.
Profilo di Oscillazione e Funzione delle Cicline
Ogni ciclina ha un profilo di concentrazione specifico che definisce il momento della sua attività:
| Ciclina | Profilo di Concentrazione | CDK partner principali | Funzione Principale |
|---|---|---|---|
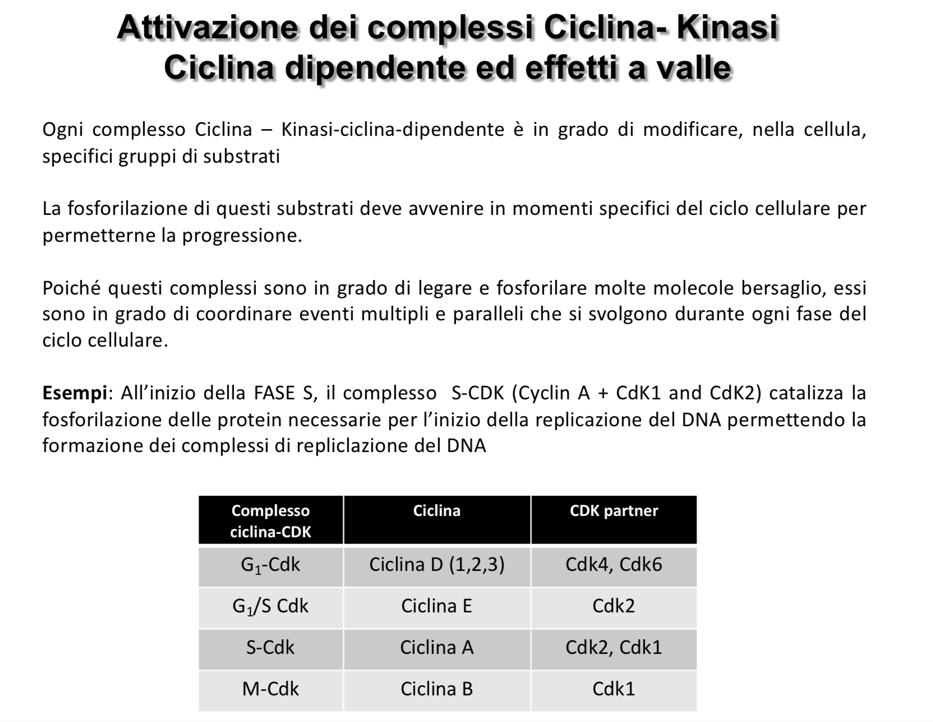
| Ciclina D | Aumenta in modo lineare durante la fase , picco tra S e . | CDK4, CDK6 | Promuove la crescita cellulare e prepara l’ingresso in fase . |
| Ciclina E | Raggiunge il picco alla transizione . | CDK2 | Permette il superamento del checkpoint e avvia la replicazione del DNA. |
| Ciclina A | Raggiunge il picco tra la fase S e la fase . | CDK1, CDK2 | Prepara la cellula e le strutture per l’avvio della mitosi. |
| Ciclina B | Raggiunge il picco durante la fase (mitosi). | CDK1 | Guida la progressione attraverso la mitosi; la sua degradazione ne determina la conclusione. |
Regolazione dell’Attività dei Complessi Cicline-CDK
Il semplice legame della ciclina alla CDK non è sufficiente per attivare pienamente la funzione chinasica. Il complesso subisce una complessa modulazione post-traduzionale:
1. Fosforilazione e Defosforilazione della CDK
Sulla struttura delle CDK sono presenti tre residui amminoacidici chiave soggetti a fosforilazione, con effetti opposti:
- Treonina 160 (): La sua fosforilazione ha un effetto attivatore sul complesso.
- Treonina 14 () e Tirosina 15 (): La loro fosforilazione ha una funzione inibitoria. Il complesso ciclina-CDK è tenuto spento finché specifiche proteine fosfatasi non rimuovono questi due residui di fosfato inibitori.
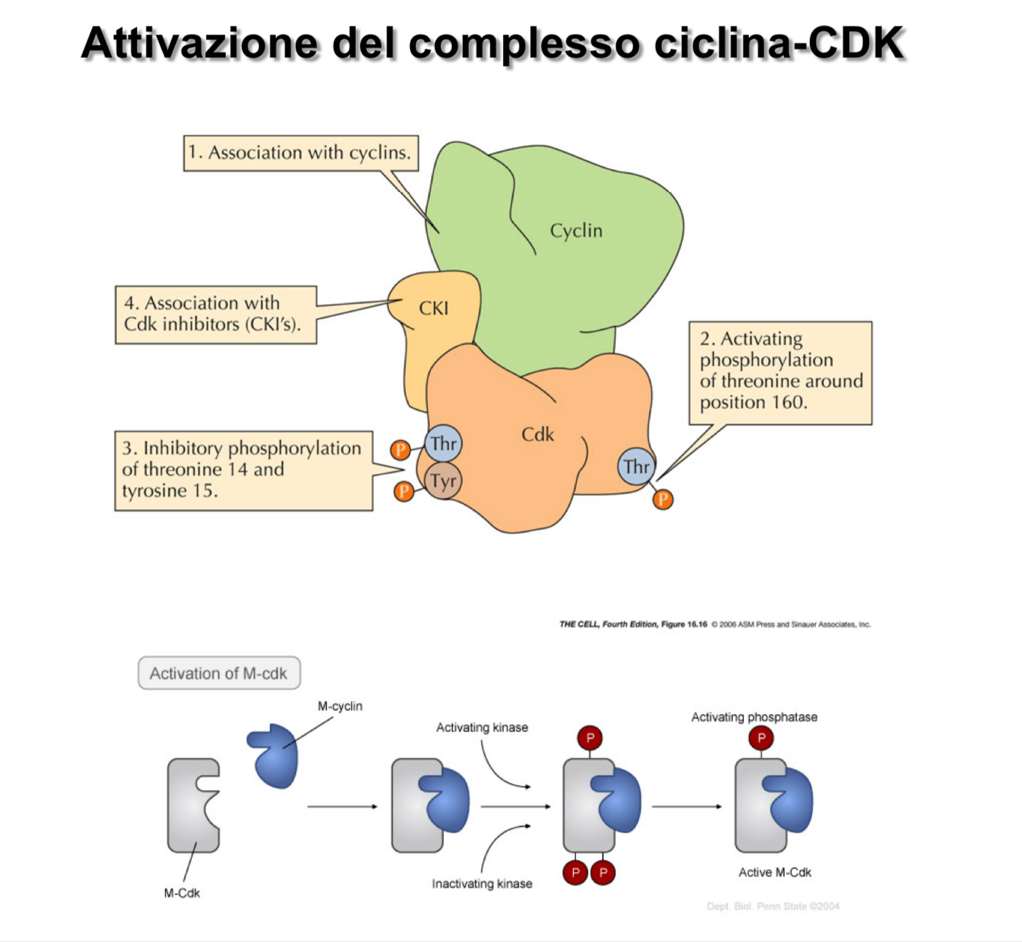
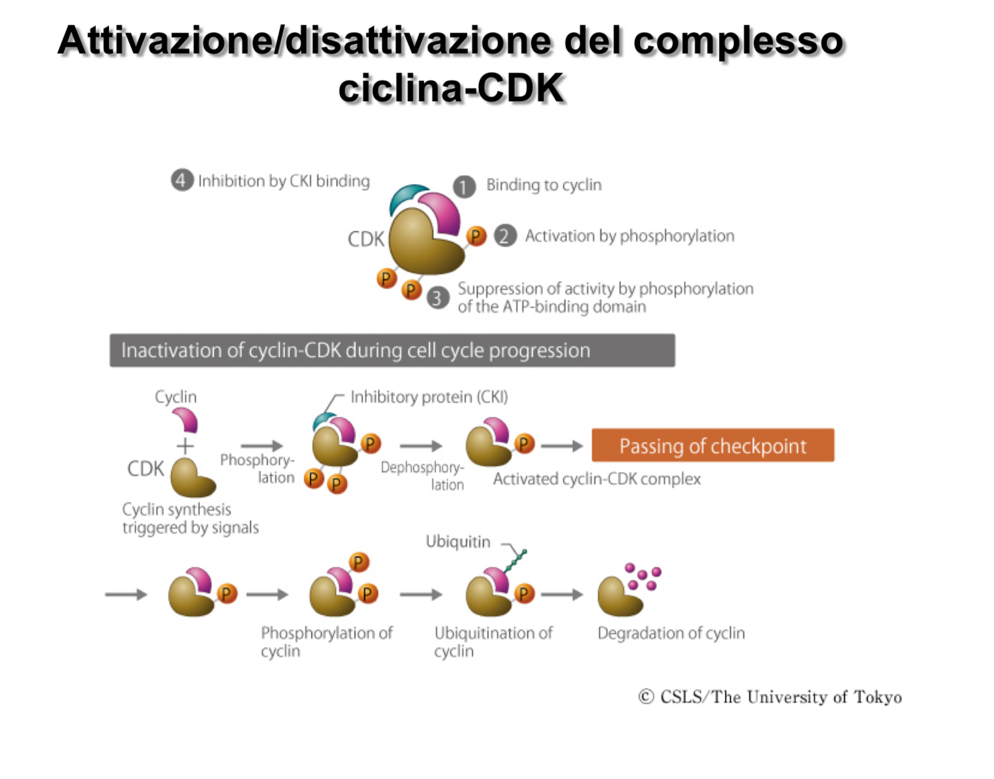
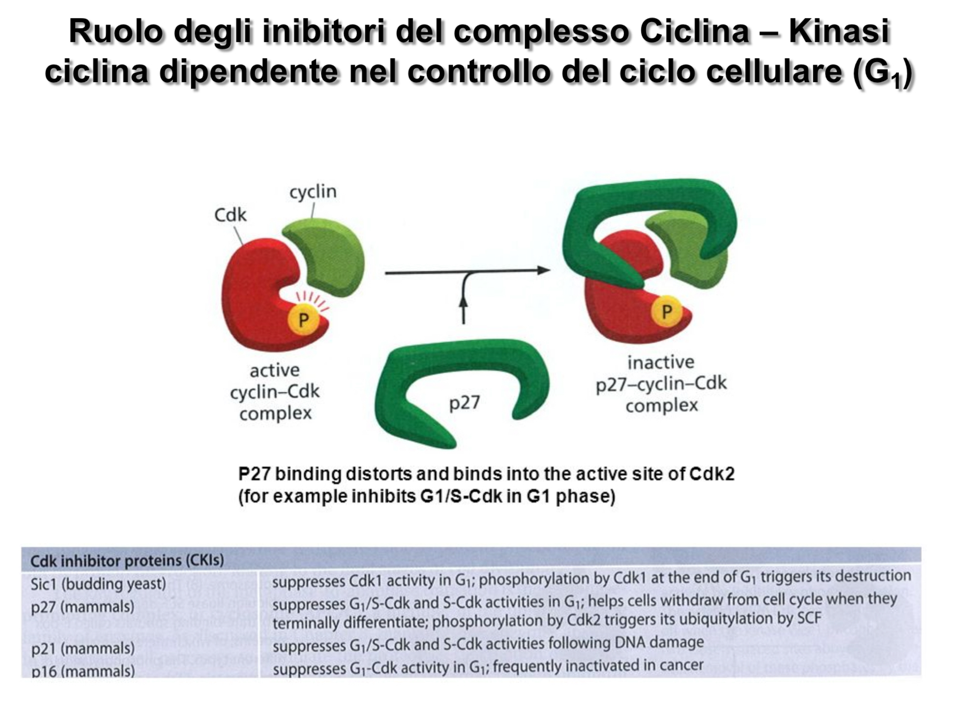
2. Inibitori delle CDK (CKI)
I complessi possono essere inattivati fisicamente dal legame con proteine regolatrici note come CKI (Inibitori delle chinasi ciclina-dipendenti).
- Meccanismo: La CKI si associa al complesso interferendo fisicamente e ostruendo il sito catalitico della CDK, impedendo la fosforilazione dei substrati.
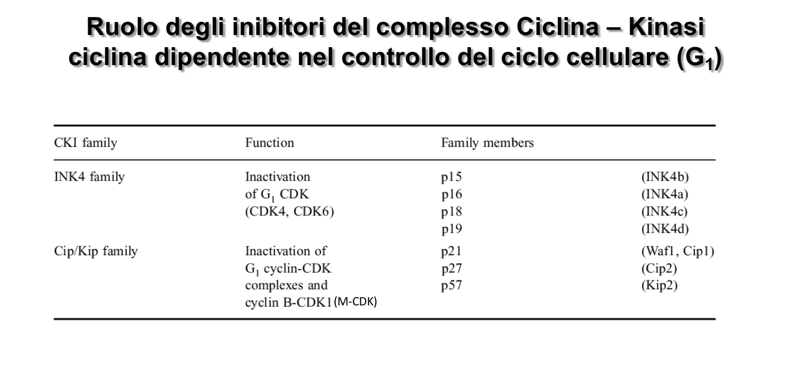
- Isoforme nei mammiferi: Le tre isoforme principali sono p27, p21 e p16 (nomenclatura basata sul peso molecolare in kDa).
Substrati dei Complessi Cicline-CDK
Ciascun complesso attivo fosforila specifici set di substrati per determinare la progressione del ciclo cellulare. Per il quadro completo complesso per complesso (compresi i complessi G1: D-CDK4/6 ed E-CDK2 → Rb) vedi Substrati delle CDK.
- Complesso Ciclina A - CDK1/2 (Inizio fase S): Catalizza la fosforilazione delle proteine necessarie alla formazione dei complessi di replicazione del DNA, avviando la duplicazione genomica.
- Complesso Ciclina B - CDK1 (Mitosi): Fosforila una serie di bersagli critici per l’ingresso e la progressione della mitosi:
- Condensina: Promuove la condensazione della cromatina in cromosomi mitotici.
- Lamine nucleari: Determina la disgregazione della rete laminare e il conseguente dissolvimento dell’involucro nucleare.
- Proteine associate ai microtubuli: Regola l’assemblaggio e l’organizzazione del fuso mitotico.
- Proteine dell’Apparato di Golgi: Promuove la frammentazione del Golgi in mini-stack.
- Separasi: Enzima che promuove la separazione dei cromatidi fratelli all’anafase tagliando le coesine residue (l’attivazione contemporanea della separasi garantisce la segregazione sincrona). Dettaglio in Mitosi (anafase).
Classificazione Oncogenetica
I geni coinvolti nel controllo del ciclo cellulare possono essere divisi in base al loro ruolo nello sviluppo tumorale:
- Proto-oncogeni: Geni che codificano per proteine che promuovono la progressione cellulare (es. i geni per le cicline e per le CDK). Se mutati in senso iperattivo (oncogeni), spingono la cellula verso la trasformazione tumorale.
- Oncosoppressori: Geni che codificano per proteine con funzione inibitoria o di controllo (es. i geni per gli inibitori p27, p21, p16 e i promotori dell’apoptosi). Mutazioni inattivanti di questi geni rimuovono i freni alla proliferazione.
(Integrazione da: sbobina 12)
🔗 Collegamenti
- Ciclo Cellulare — 📋 fa parte di
- Sistemi di Controllo del Ciclo Cellulare — 🔗 stesso meccanismo
- Enzimi — 📋 fa parte di
- Mitosi — ➡️ evoluzione
- Oncogenesi — 🔄 connessione clinica
- Modificazioni Post-Traduzionali — 🔗 stesso meccanismo (ubiquitina–proteasoma)
- Regolazione Extracellulare del Ciclo Cellulare — ⬆️ causa (mitogeni → Ciclina D)
- Substrati delle CDK — 🔍 approfondimento (substrati per ogni complesso)